

What is the future for The Martes Complex (Guloninae) in the face of climate change and ecological breakdown?
2 Vincent Wildlife Trust, Ledbury, HR8 1EP, UK
3 NatureSpy, Mildenhall, IP28 7DE, UK
JM: https://orcid.org/0000-0002-6919-120X

Abstract photo. European pine marten. Photo credit David Baird.
Abstract
The climate and ecological emergency (CEE) is impacting the natural world with increasing severity, with likely serious consequences for the Martes Complex (Guloninae) including range contraction and fragmentation, ecosystem collapse and disrupted inter-species relationships. We assessed the relative vulnerability of each species of Guloninae to the CEE based on IUCN listings, threats to preferred habitat, habitat specificity, global range size, and the nature and number of Köppen-Geiger climate zones occupied. This assessment indicated that ten of the eleven Guloninae are moderately or highly vulnerable to the impacts of the CEE, and only the stone marten Martes foina is of relatively low vulnerability. We explored biotic and abiotic factors to identify the range of known and likely impacts upon Guloninae, with deforestation and wildfires as recurring themes. Mitigation for Guloninae should focus on landscape-scale efforts to expand and connect suitable forested habitat, ideally on a north-south axis so as to facilitate predicted range shifts. In anticipation of reduced funding for wildlife research and conservation in an increasingly unstable geopolitical context, the best opportunities for mitigation will likely depend upon influencing the forestry industry and national carbon capture reforestation policies.
Keywords: biodiversity loss, climate change, global heating, Guloninae, Martes, mitigation, reforestation
Introduction
The Earth’s life-support systems are breaking down in response to global-scale processes driven by human activity. Referred to as the Climate and Ecological Emergency (CEE) (Gardner et al., 2021) because of the prominent influence of anthropogenic global heating and biodiversity loss, its impacts are increasingly harming humanity and the natural world (Fletcher et al., 2024). Notably, as a consequence of rising emissions of greenhouse gasses such as carbon dioxide, nitrous oxide and methane, the climatic stability that characterized much of the Holocene has been replaced by increasing global temperatures, rising sea levels, unpredictable seasons and a growing frequency and intensity of extreme weather events such as heatwaves, droughts, wildfires, high-energy storms, torrential rain and severe flooding (IPCC, 2023). Some scientists argue that we have left the Holocene and entered the Anthropocene, a new geological epoch characterized by the scale and severity of planetary processes driven by human activity (Zalasiewicz et al., 2010). Increasingly scientists view climate breakdown and biodiversity loss as symptoms of ecological overshoot driven by a crisis of unsustainable human behavior (Merz et al., 2023; Fletcher et al., 2024).
Since 1970 global populations of vertebrate wildlife have fallen on average by more than two-thirds (Almond et al., 2020). With an estimated one million species at risk of extinction, the five key drivers of biodiversity loss identified by the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES, 2019) are: changes in land and sea use; direct exploitation of natural resources; climate change; pollution; and invasion of alien species. Ceballos and Ehrlich (2023) warn that we are in a human-driven sixth mass extinction event that is more severe than previously assessed and rapidly accelerating; predicted extinction rates for mammals are especially high at 511 times above background rates.
One of many consequences of the CEE is that ecosystems around the globe are being impacted by unprecedented environmental conditions that now compromise their capacity to support endemic species. The high probability of ever more catastrophic ecosystem impacts requires biologists to focus on identifying and mitigating impacts and helping natural systems to adapt to change as far and as fast as this is possible (Lawler et al., 2024). Whilst the CEE is a consequence of several anthropogenic drivers (IPBES, 2019), current societal concern and associated research effort relates primarily to the impacts of global heating and climate change. This is reflected in the focus of this paper.
In their review of efforts to assess species’ vulnerability to climate change, Pacifici et al. (2015) defined vulnerability as ‘the predisposition to be adversely affected’, with both intrinsic and extrinsic factors contributing. Assessing the vulnerability of species to climate change typically involves an evaluation of exposure (the type and magnitude of climatic changes that the species has experienced or is projected to experience), sensitivity (how much it is affected by or susceptible to a climate-related change) and adaptive capacity (the species’ ability to cope with, adjust to and persist under current and future climate change) (Thurman et al., 2020). Attempts at predicting the likely effects of climate change on species have used a range of different measures to assess their vulnerability, but very few (c. 4%) of the published studies reviewed by Pacifici et al. (2015) assessed species’ vulnerability globally. An important role for conservation biologists is to identify those species that are most vulnerable to environmental changes of this magnitude in order to minimize global biodiversity loss (Pacifici et al., 2015).
The purpose of this paper is to assess the relative vulnerability of Guloninae to the CEE across their global distribution and to identify the range of known and likely impacts. The Guloninae comprises the eleven species within the genera Martes, Pekania, Gulo and Eira. These species share many life history traits, habitat associations and conservation challenges as detailed by Proulx & Aubry (2017).
The Guloninae’s responses to CEE impacts are likely to vary between species according to their vulnerability and resilience, and some responses will be more predictable than others. Species’ responses may affect many aspects of their biology including population density, demography and interspecific relations (Wereszczuk, 2023). Separate from climate change impacts, a related consideration is the vulnerability of each species to the existing direct and indirect impacts of human activity upon biodiversity, such as resource use and pollution. Species that are already declining due to non-climate change-related pressures are likely to be more sensitive to climate change. For example, Pearson et al. (2014) found that decreasing population size and/or occupied area, as well as increasing range fragmentation, were associated with higher extinction risk under climate change. Degradation and loss of habitat have already been identified as major threats to many Guloninae (Proulx et al., 2004), and Carroll (2007) identifies that this threat is exacerbated by the interacting effect of climate change leading to range fragmentation.
Most Guloninae have a strong affinity for or dependence upon forested habitats (Proulx & Aubry, 2017; Wright et al., 2022). From 2001 to 2023, there was a total of 488 Mha of tree cover loss globally, equivalent to a 12% decrease in tree cover since 2000 (Global Forest Review, 2024a) and forests are under accelerating multiple pressures, including increasing resource use, wildfires, shifting agriculture, and emerging diseases (Global Forest Review, 2024a; Tew et al., 2023). The geography and chronology of forest loss vary across the globe, with many countries in the global north having undergone major deforestation some centuries ago. Rates of tree cover loss have been rising since the year 2000, and there have been consistent differences in the extent of tree cover loss across the world’s ecozones, with tropical and subtropical forests suffering greater losses than boreal and temperate forests (Global Forest Review, 2024a).
Direct anthropogenic deforestation to create space for agriculture and other land uses in boreal and temperate forests has now been superseded by increasing losses of tree cover to severe fires and insect pests, both fueled by global heating and drought related to the CEE, in addition to timber harvesting (Jolly et al., 2015; Pureswan et al., 2018). Analyses by Global Forest Watch distinguish between these temporary impacts (identified as tree cover loss) that now account for 99% of losses in boreal and temperate forests, and permanent forest clearance for agriculture and urbanization (identified as deforestation) that dominates losses in the tropics (Global Forest Review, 2024a). Currently >96% of global deforestation (as opposed to tree cover loss) occurs in the tropics (Global Forest Review, 2024a). 32 million hectares of primary or recovering forest were lost in the highly biodiverse tropics between 2010 and 2015 (IBPES, 2019). Global Forest Watch reports that tropical primary forest loss remains on an increasing trend, with 3.7 million hectares destroyed in 2023 (Global Forest Review, 2024b) and this loss must be viewed as a significant driver of harm to the tropical Guloninae.
Rising global temperatures are predicted to be one of the greatest drivers of faunal change this century (Lawler et al., 2009). A review by Pigot et al. (2023) of geographical data for approximately 36,000 wild animal species, including >5,000 mammals, indicates that global heating is already driving widespread population change and geographical range contractions. Projections to the year 2100 under different heating scenarios show that species will suffer abrupt increases in the area of their geographical ranges where they will be at risk of thermal exposure. Some Guloninae are cold-adapted, so are likely to be adversely affected by global processes driving rising temperatures and associated reductions in snow cover. For example, for cold-adapted Guloninae it is of concern that the Paris Accord’s ‘safe guardrail’ heating limit of 1.5°C (Schleussner et al., 2022) was breached in 2024 (Copernicus Climate Change Service, 2025) and large parts of the northern hemisphere were more than 1.5 degrees warmer than the 1991-2020 average (Figure 1; Copernicus Climate Change Service, 2025); and the Arctic region has been warming nearly four times faster than the global average since 1979 (Rantanen et al., 2022). Current policies intended to curb global emissions of greenhouse gases are inadequate and average temperatures are projected to increase to 2.7°C above pre-industrial levels by 2100 (Climate Action Tracker, 2023).
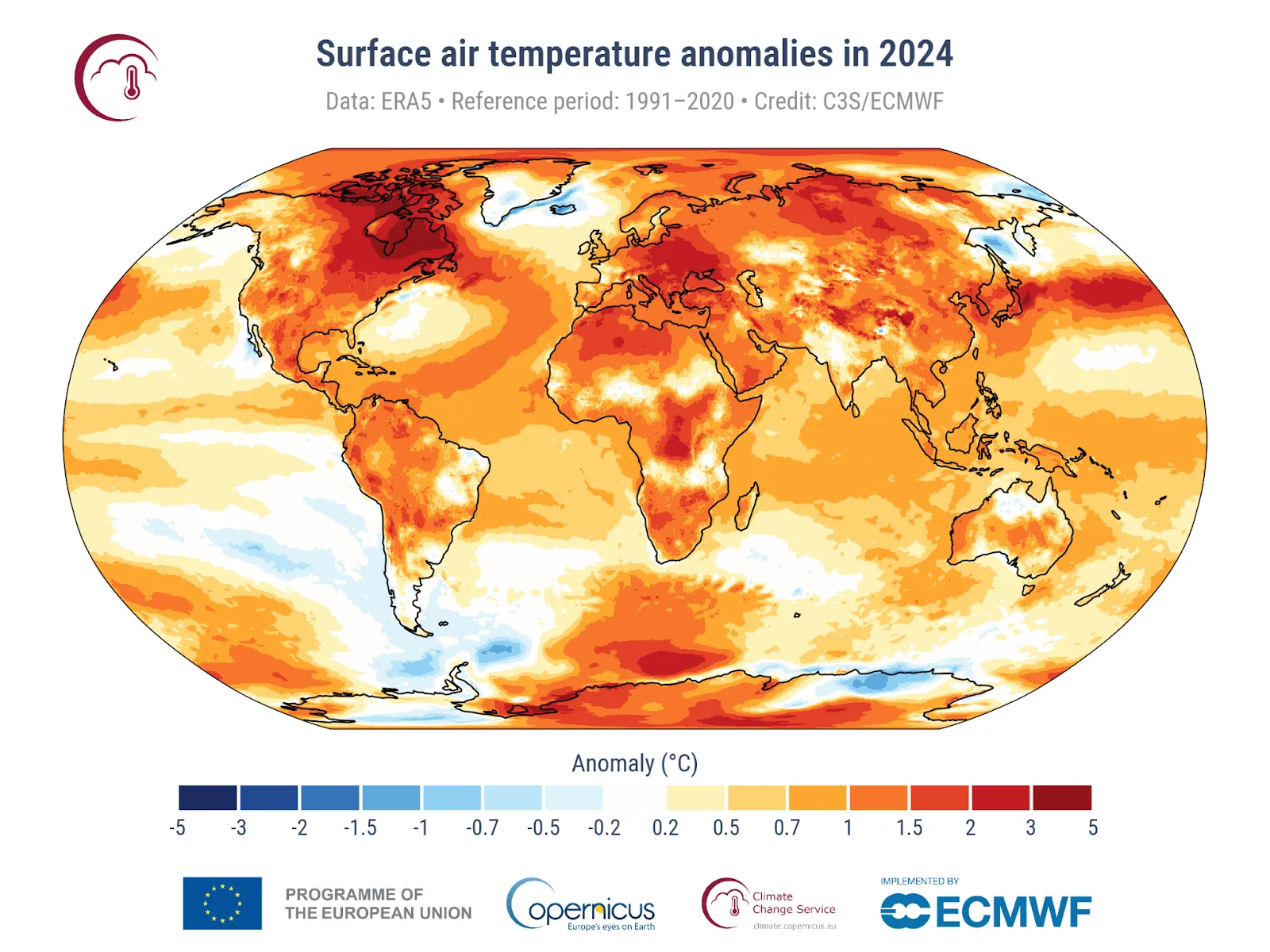
Figure 1. Average surface air temperature anomaly in 2024 compared with the 1991-2020 average, showing that most of the world was hotter than normal in the record-breaking year of 2024. Notably, in 2024 parts of North America and Eurasia occupied by cold-adapted members of the Guloninae were more than 1.5°C warmer than the 1991-2020 average (Copernicus Climate Change Service, 2025).
Methods and Materials
Biological sampling
To assess the relative vulnerability of Guloninae species to the CEE we used a trait-based vulnerability assessment (TVA) framework. This framework uses the three dimensions of climate change vulnerability, namely sensitivity, exposure and adaptive capacity, to assign species to vulnerability classes (Foden et al., 2013). Although TVAs such as ours that are based on expert opinion and interpretation of the published literature can be somewhat subjective, the method nevertheless allows for a relatively rapid assessment of multiple species, which is useful for prioritizing and implementing conservation measures and is accessible to practitioners who may not have extensive knowledge of modeling techniques.
Each of the eleven Guloninae species (see Figure 2) was assigned – via the process outlined below – to a high, medium or low overall relative vulnerability class based on their combination of sensitivity, exposure and adaptive capacity. Species identified as having high vulnerability to the CEE are of greatest concern and are the highest priority for monitoring responses to climate change and likely the most at risk of serious harm in the absence of effective conservation action. Species identified as having medium vulnerability to the CEE have the potential to adapt but warrant close monitoring. Species identified as having low vulnerability to the CEE are those most likely to withstand harmful impacts but monitoring is needed to ensure that this assessment of vulnerability is realized.
We assessed seven factors to demonstrate sensitivity, exposure and adaptive capacity and used these to assign each species to a class of relative vulnerability to the CEE:
- Global range size
- Number of Köppen-Geiger climate zones (Peel et al., 2007; Beck et al., 2018) occupied
- Nature of Köppen-Geiger climate zones occupied
- Level of threat to preferred habitat
- Habitat specificity
- IUCN listing
- IUCN population trend
We accept that some of these characters overlap and interact to some degree. Nevertheless, we believe they provide a robust basis upon which to derive for the Guloninae a three-way classification of relative vulnerability to the CEE. We explain our reasoning below.
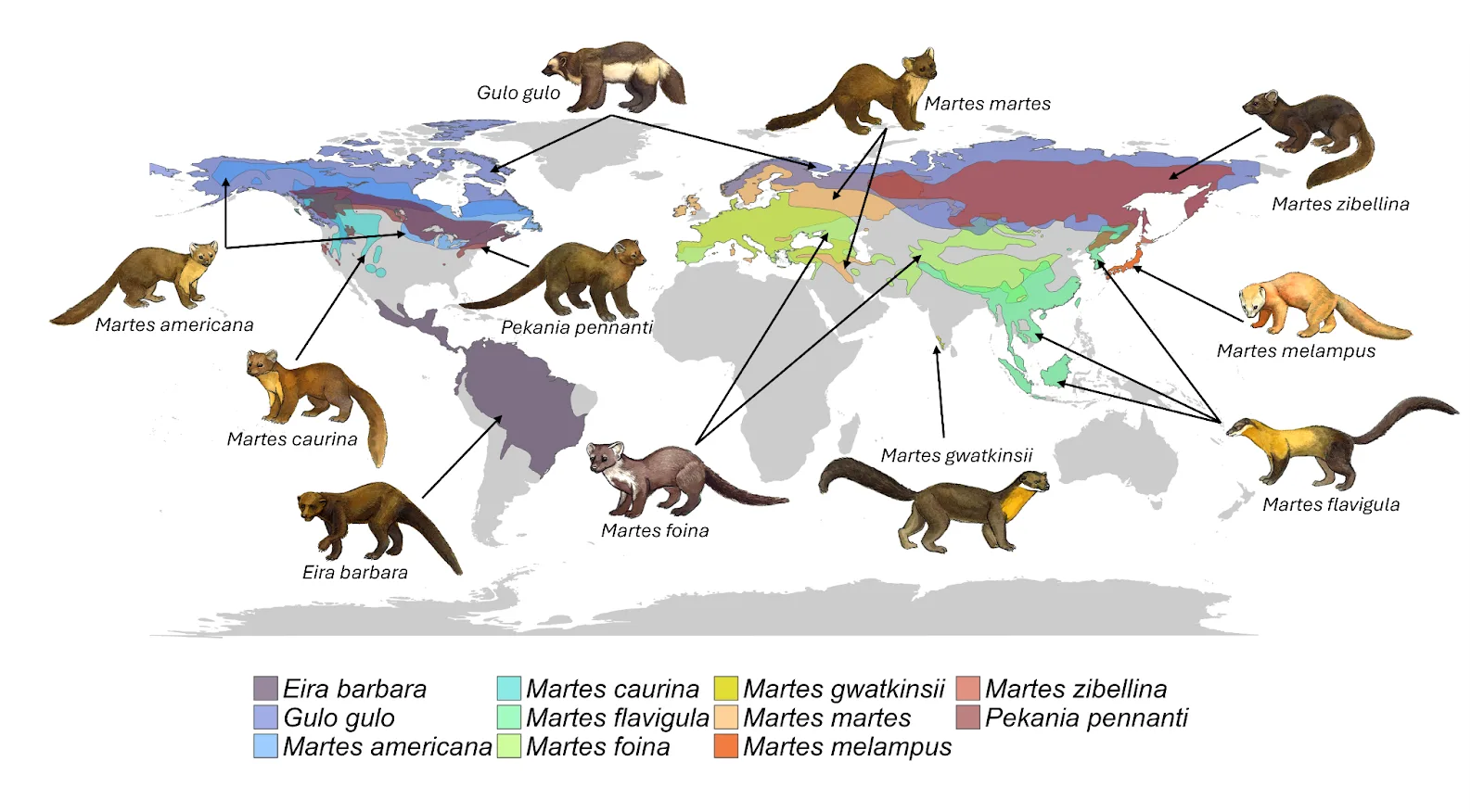
Figure 2. Global distribution of the eleven Guloninae species (Burgar et al., 2025), reproduced with permission from the authors. For up-to-date ranges, refer to the Martes Working Group website www.martesworkinggroup.org.
Global range size
We took the view that species with large global ranges were likely more resilient to CEE impacts than those with small global ranges. Global range location and size of maximum range extent in square kilometers were derived from the most recent IUCN listings. These represent the limits of the current known distribution from occurrence data along with knowledge of habitat, elevation limits and expert knowledge of the species and its range. However, the polygons displayed on the distribution maps communicate that the species probably only occurs within the polygon but it is not necessarily distributed equally nor does it occur everywhere within that polygon (IUCN, 2018). We categorized global range sizes of Guloninae as Small (<1 million km2), Medium (1-10 million km
Number of Köppen-Geiger climate zones occupied
We used the Köppen-Geiger system (Peel et al., 2007; Beck et al., 2018) that classifies global climate into five main zones: Tropical; Arid; Temperate; Continental; and Polar (Figure 3). We compared the global range of each species with the distribution of the five main climate zones to identify the nature and number of zones they currently occupy. We assumed that those species confined to just one climate zone would be more sensitive to climate change than those occupying two or more zones because of their lower intrinsic adaptive capacity and their inability to shift their range across multiple climate zones. Accordingly we chose the following categories: one climate zone occupied = High sensitivity; two climate zones occupied = Medium; and three climate zones occupied = Low.
Nature of Köppen-Geiger climate zones occupied
In light of evidence of regional differences in current rates of global heating and the strong association of cold-adapted Guloninae species with the faster-warming Polar and Continental climate zones, we assumed that there are differences in their vulnerability to global heating linked to the nature of the zone that each species predominantly occupies as follows: range predominantly in the Polar zone = High vulnerability; range predominantly in the Continental zone = Medium; range not in the Polar or Continental zones = Low.
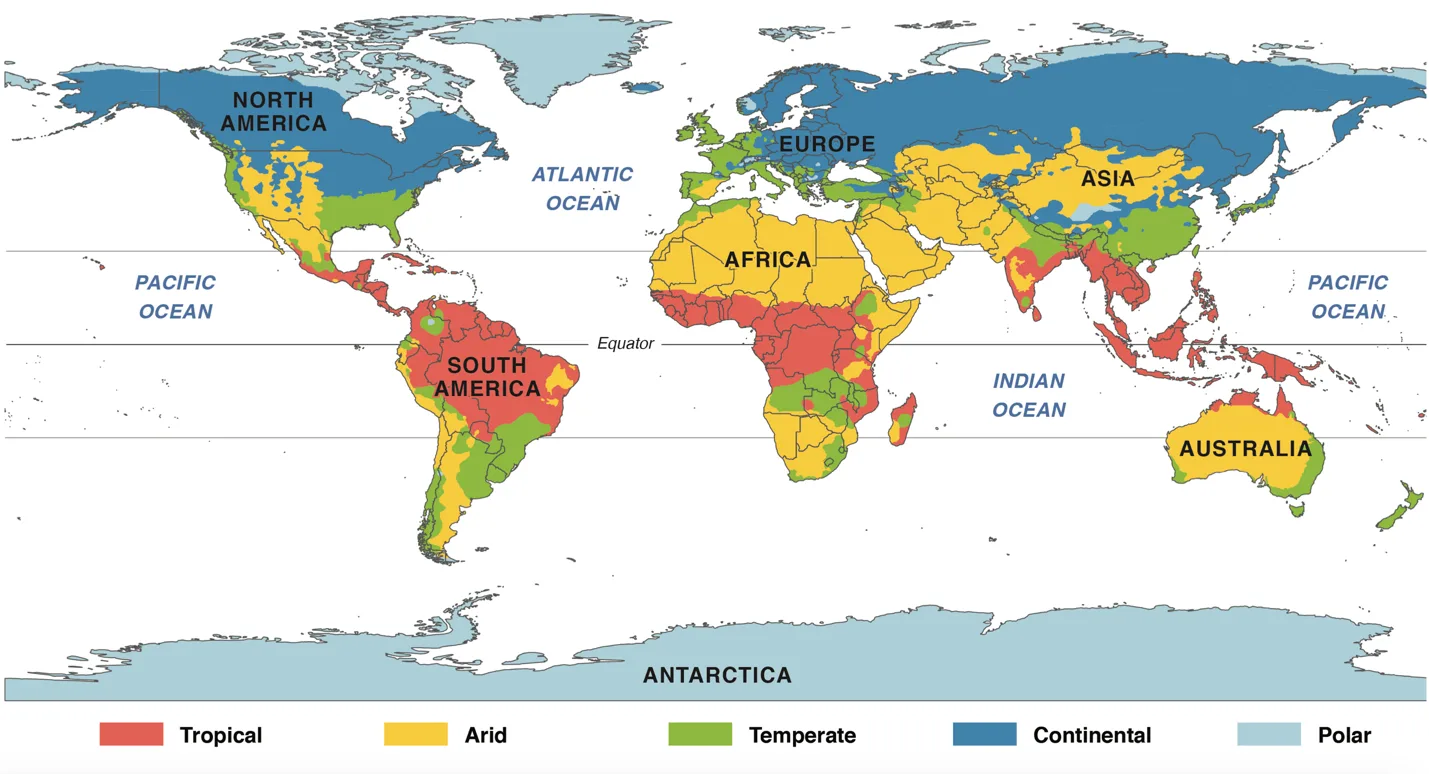
Figure 3. The current global distribution of the five main climate zones defined and described by the Köppen-Geiger system (Peel et al., 2007; Beck et al., 2018). Image credit: Emily Eng, Smithsonian Institution https://ssec.si.edu/sites/default/files/other/climate/ClimateZoneMap-.pdf.
Level of threat to preferred habitat
In light of evidence of marked differences in current rates of forest loss between the tropics and other bioclimatic zones we assigned a level of exposure to the CEE based on the level of threat to the forested habitat that each species occupies as follows: High = species occupying tropical forests; Medium = species occupying non-tropical forests; Low = species showing no strong preference for occupying forested habitat. We derived information on forest occupancy for each of the eleven species from a combination of expert opinion and interpretation of the current literature.
Habitat specificity
The degree of habitat specificity for the Guloninae species was also derived from expert opinion and interpretation of the current literature. We used the habitat specificity of each species as a measure of their adaptability to changing conditions in their preferred habitat as follows: High = species rarely recorded outside their preferred habitat demonstrating low adaptability; Medium = species demonstrating tolerance of moving through a heterogeneous habitat matrix; and Low = species recorded throughout heterogeneous habitat outside of their preferred habitat demonstrating high adaptability. Species identified as having a high degree of habitat specificity were deemed to be less adaptive and therefore have a higher vulnerability to the CEE than those demonstrating greater adaptive capacity with a medium or low degree of habitat specificity.
IUCN listing
We recognized that current IUCN listings may not all be accurate because the latest assessment was completed in 2015. For example, it does not separate the genetically distinct Pacific marten M. caurina and American marten M. americana (Carr & Hicks, 1997; Dawson et al., 2012). We assumed that those species listed by IUCN as ‘Endangered’ are the most sensitive to the CEE (= High sensitivity) followed by those that are listed as ‘Vulnerable’ (= Medium) and as ‘Least Concern’ (= Low) respectively.
IUCN population trend
We considered the IUCN population trends of each of the eleven species to assess their current sensitivity to the CEE. Informed by Pearson et al. (2014), we assumed that species whose populations are already decreasing as a result of existing and historical threats (such as habitat loss, over harvesting and non-native invasives) will be more sensitive to the impacts of the CEE than those species that are currently not experiencing decline as a result of pre-existing threats. Accordingly, we assumed that those species with a ‘Decreasing’ population trend are more sensitive to the CEE (= High sensitivity) than those with a ‘Stable’ (= Medium) or ‘Increasing’ (= Low) population trend respectively.
Having categorized and/or quantified the factors as explained above, we identified the contribution each factor made to the overall relative vulnerability of species to the CEE as shown in Table 1. This process involved a quantification of overall vulnerability based upon the total scores for each species, where High factors scored 3, Medium factors scored 2 and Low factors scored 1.
In order to define overall relative vulnerability based upon the sum of the seven factor threshold scores we used the following thresholds:
- Low overall vulnerability (9-11)
- Medium overall vulnerability (12-14)
- High overall vulnerability (15-17)
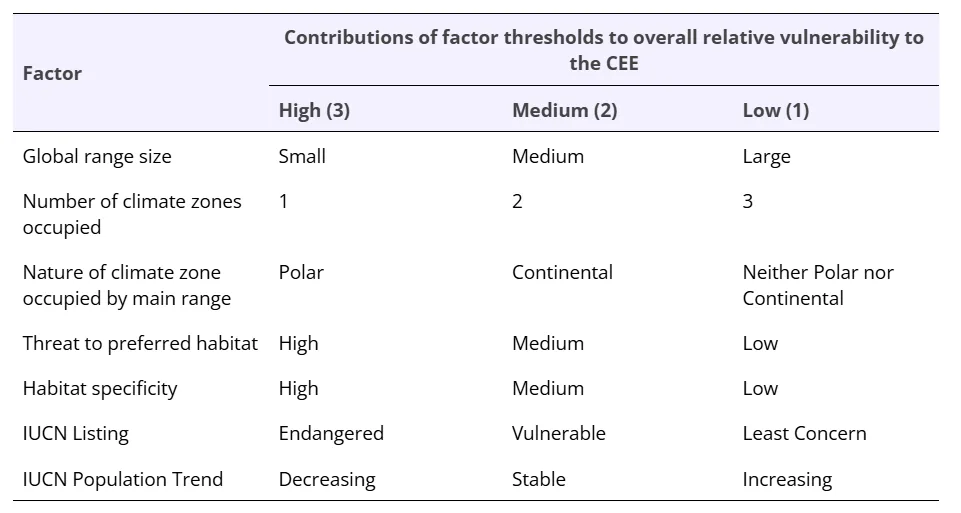
Table 1. A summary of the three-way categorisation of each of seven factors assumed to define relative vulnerability to the Climate and Ecological Emergency (CEE). Species are classified as having High, Medium, or Low overall vulnerability to the CEE based on the scored thresholds of the seven contributing factors.
Results
Details of the contribution of the seven factors to the overall vulnerability to the CEE of the eleven Guloninae species and their overall relative vulnerability score are presented in Table 2. Three of the eleven Guloninae species were assessed as having high overall vulnerability to the CEE, comprising the American, Pacific and Nilgiri martens. Six species were assessed as having medium overall vulnerability to the CEE: Sable; yellow-throated marten; Japanese marten; pine marten; fisher; wolverine and tayra. Only the stone marten was assessed as being of relatively low overall vulnerability to the CEE.
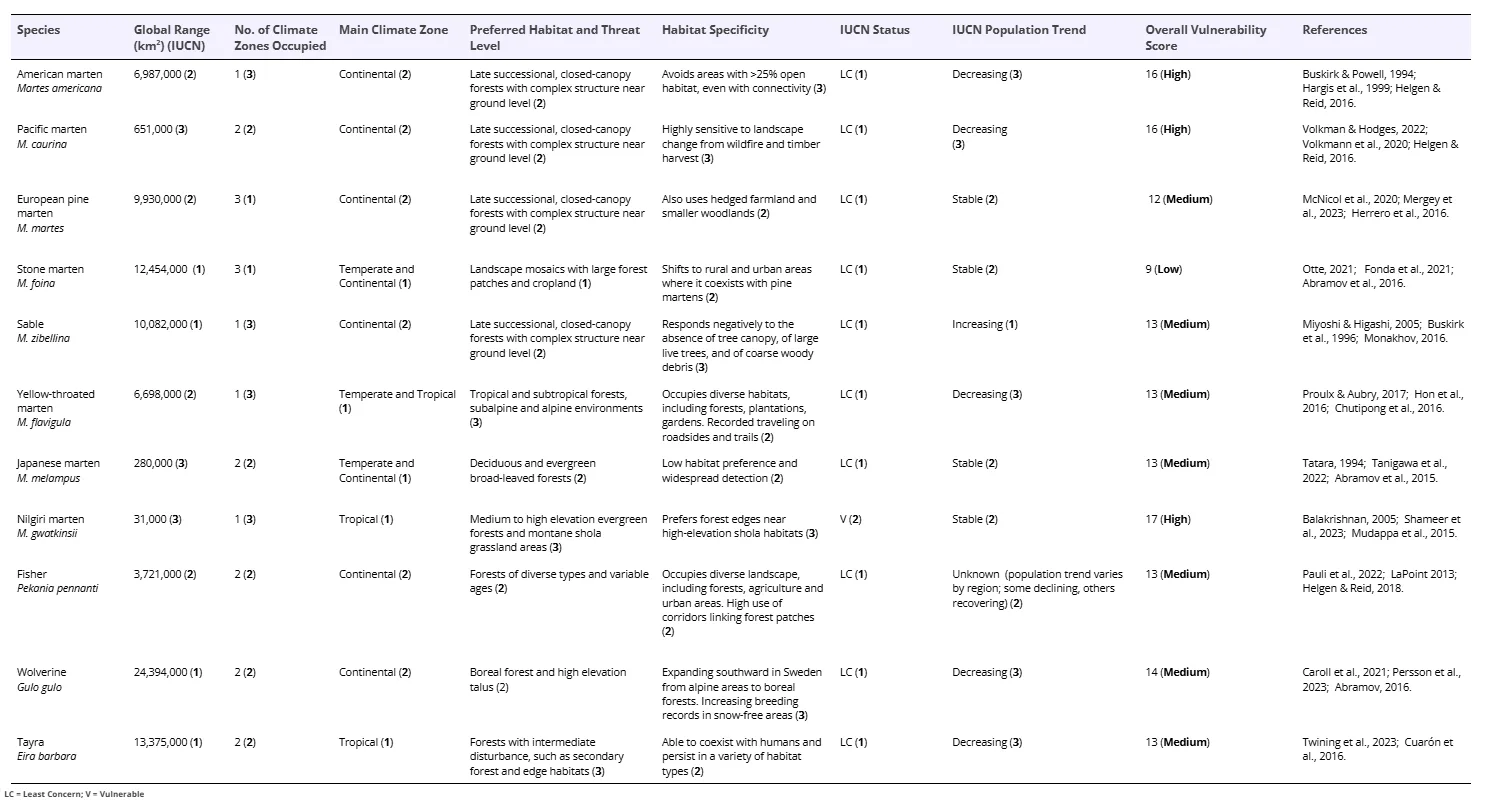
Table 2. A summary of the contribution of each of the seven factors to the overall vulnerability of the 11 Guloninae species to the Climate and Ecological Emergency (CEE) based on the threshold classifications described above. Relative vulnerability scores are shown as bold numbers (see Table 1 for definitions). Note that the threat level number in the preferred habitat column relates to the threat to that habitat (see Table 1). Note that the IUCN gives no population trend for Fisher, so we have assigned this factor a ‘Medium’ score. Also, because the IUCN does not separate the Pacific marten from the American marten, we have assigned the same IUCN factor scores to each of these species.
Direct impacts of climate change on Guloninae
Whilst our assessment indicates that most of the Guloninae are moderately or highly vulnerable to the CEE, some impacts of rising temperatures are difficult to predict in the absence of data on the thermal niche of each species. Nevertheless, the knowledge that eight of the 11 Guloninae species are cold-adapted and associated with the Polar and/or Continental Köppen-Geiger climate zones, together with evidence that the Arctic is warming four times faster than the global average (Rantanen et al., 2022), suggests that these species may already be experiencing significant effects.
Increasing temperatures will likely drive the northward and upslope range contraction of cold-adapted species; this may induce increased range fragmentation and isolation of sub-populations given limited connectivity of suitable habitat, and potential risks of increased intra- or inter-specific conflict (Elsen & Tingley, 2015; Vergara et al., 2016). Where more thermophilic species are sympatric, such range shifts may involve the replacement of one species by another. For example, modeling of Guloninae responses in Europe have predicted northward range contraction of the cold-adapted pine marten and northward range expansion by the more habitat-generalist and adaptable stone marten (Vergara et al., 2016; Lawler et al., 2012). Similarly, modeling predictions by Spencer et al. (2015) indicate that Pacific marten in the Sierra Nevada is likely to suffer from habitat loss and fragmentation under climate change, with upslope shrinkage of habitat; in contrast, future predictions for fisher varied between climate scenarios, with some predicting habitat loss and others habitat gain. Zielinski et al. (2017) affirm the uncertainty of outcomes where responses to climate change of competing Guloninae at their range edge are difficult to predict.
An empirical example of range shift response by a cold-adapted Guloninae is reported in Hiltner’s (2022) study of the American marten in Michigan’s Upper Peninsula, at the southern edge of the species’ US range where an earlier study by Lawler et al. (2012) predicted that climate change would shrink and degrade marten habitat. Harvest data between 2005 and 2020 revealed a gradual decrease in marten occupancy in the southern part of the study area in response to a decline in the area of suitable abiotic conditions, notably deep snow and low mean winter temperatures (Hiltner, 2022). In light of wider predictions of northward range shifts by this species in response to global heating, Hiltner (2022) argues that an understanding of the drivers of change at the southern border will better inform conservation action for American martens. A separate modeling study of American martens in the northern Appalachians (Carroll, 2007) predicted a 40% population decline by 2055 in response to reduced snow cover due to global heating.
Impacts of wildfires on Guloninae
Outside of the tropics, global trends in tree cover loss are driven by fire dynamics in boreal forests (Global Forest Review, 2024a). These events rapidly damage or destroy large areas of habitat, with a variety of context-dependent responses from the carnivores that occupy them (Geary et al., 2020). The frequency, extent and severity of wildfire events are increasing in response to global heating and associated drought conditions (IPCC, 2023; Jolly et al., 2015). For example, 2023 was remarkable for the extent of forest destroyed by wildfires across many countries, damaging the habitat of several northern Guloninae species. Notably, Canada had its worst wildfire season on record, with a fivefold increase in tree cover loss due to fire compared with 2022 (Global Forest Review, 2024b). Research studies on the impacts of changing fire regimes upon carnivores occupying boreal and montane forests in Canada and the USA are undermined by taxonomic bias and inconsistent approaches (review by Volkmann et al., 2020).
Further south, increasingly severe wildfires have impacted the habitat of fisher and Pacific marten in the western USA: movements of Pacific martens revealed complex responses to post-fire forest landscapes, with extreme avoidance of areas subject to salvage logging, the practice of extracting trees from disturbed areas to minimize loss of commercial timber (Volkmann & Hodges, 2021). A study of fishers – a species of conservation concern in the western U.S. – by Green et al. (2022) revealed decreases in fisher abundance following damage to habitat in the Sierra Nevada caused by wildfires and post-fire forest management such as salvage logging; the authors concluded that the increasing severity, frequency and extent of wildfires in the western USA will harm fisher populations and alter the composition of mesocarnivore communities.
Biotic factors driving changes in Guloninae behavior, range and/or abundance
Biotic responses to the CEE will influence the structure and function of ecosystems and their capacity to support healthy populations of Guloninae species. Impacts may range between minor shifts in the availability of resources, to complete ecosystem breakdown. ‘Catastrophic forest ecosystem collapse’ was the highest ranked issue among a majority (64%) of Expert Panel members considering the future of UK forests over the next 50 years (Tew et al., 2023). Inevitably, catastrophic consequences of global climate breakdown for forest habitats will be widespread beyond the UK, with serious impacts upon tree health and forest structure arising from the increasing intensity and frequency of drought, wildfires and storms (IPCC, 2023). The rising impact of pests such as bark beetles on trees rendered more vulnerable by climatic stress has led to predictions of wide scale collapse of boreal forests – important Guloninae habitat and a major global carbon sink – across the northern hemisphere (Greenfield, 2024).
Predicting outcomes for Guloninae is challenging because of the complexity of ecosystem components, interactions and processes involved. Nevertheless, identifying the drivers that stress and threaten populations in an era of rapid environmental change is crucial (Kuntze et al., 2024). The examples below offer some insights into the scale of that challenge.
Some climate-related impacts upon Guloninae are driven by prey populations responding to rapid environmental change. Shifts in prey abundance, distribution and the timing of activity (Saino et al., 2011; Schweiger et al., 2008) may have profound impacts upon predators. For example, in the Sierra Nevada of California wide-scale climate-induced tree mortality induced a dietary shift from mammals to plants in an endangered population of fishers because of reductions in the availability of preferred mammalian prey (two species of tree squirrel), with likely consequences for the behavior, energetics, demography and fitness of the population (Pilgrim et al., 2023). Other studies have demonstrated effects of tree mortality on physiology, behavior and survival of fishers (Kordosky et al., 2021a; Kordosky et al., 2021b; Kuntze et al., 2024). Changes in forest composition and snow cover regimes have also resulted in northward shifts in important prey populations, with possible implications for American marten populations (Scott et al., 2022).
Drought-induced tree mortality was found to be the main driver of stress levels in fishers in the California population, with associated demographic impacts including reduced survival rates (Kordosky et al., 2021a; Kuntze et al., 2024). The survival of this isolated fisher population at the southern end of the Sierra Nevada mountain range is now uncertain because of the impacts of climate change, including the rapid increase in the size and severity of wildfires driving habitat loss and fragmentation (W. Spencer, pers. comm. [paper presented to Aviemore Martes Symposium]; Spencer et al., 2015; Kuntze et al., 2024; Steel et al., 2023).
Differential responses of two North American Guloninae to climate change and forest management have been revealed by trapper interviews in eastern Canada (Suffice et al., 2017): fishers had increased and expanded their range since the early 2000s, while American martens had decreased. Results from interviews suggest that forest management and climate change conferred localized benefits upon fishers, while negatively influencing American martens due to declining habitat and increased predation and competition from fishers and other carnivores. An earlier study highlighted the role of snow in segregating American martens and fishers (Karniski, 2014). Given the similarity between these two species, increased agonistic interactions driven by reduced snow cover are likely to be detrimental to the smaller American martens, which are occasionally killed by fishers (McCann et al., 2010).
In Poland, a global heating-induced increase in primary productivity and woodland rodent populations is believed to have driven a change in skull morphology and increase in pine marten body size since 1980, together with a shift towards a male-biased sex ratio over the same period (Wereszczuk et al., 2023). Moreover, a wider study of both pine and stone martens across continental Europe revealed differential rates of climate warming-induced body mass increase (over the period 1960-2019 pine marten average body mass increased by 24%; stone martens by 6%), with stronger interspecific competition a potential consequence in areas of sympatry (Wereszczuk et al., 2021). These are part of a suite of unpredictable morphological changes reported across many taxa as species respond to new conditions delivered by global heating (Goldenberg et al., 2022; Weeks et al., 2020).
Socio-economic impacts upon Guloninae research and conservation
As human societies around the globe experience the increasingly harmful impacts of the CEE, the likelihood of energy, food and water insecurity, mass migration, international conflict and societal breakdown will grow (Richards et al., 2021). In response, national governments will necessarily change policies and funding priorities in their efforts to avert or minimize economic and societal collapse. Funding for wildlife research and conservation will likely shrink to the extent that most work on Guloninae will lose financial support.
Under a CEE scenario of public funds shifting away from Guloninae conservation, the continuation of such work will depend upon the extent to which higher priority policy areas might be influenced to benefit Guloninae. For example, reforestation is now prioritized by governments because of its role in carbon sequestration (e.g. Forestry Commission, 2022; Cho et al., 2025) and the supply of carbon neutral material for the construction industry (Atkinson et al., 2022; Rapid Transition Alliance, 2021). A further incentive for extensive tree planting arises from new evidence that afforestation contributes to local air cooling and climate mitigation (Barnes et al., 2024). Herein lie opportunities to influence the scale and pattern of afforestation and reforestation to mitigate CEE impacts upon Guloninae (see below).
Mitigating the CEE’s impacts upon Guloninae
The collective global failure to address the causes of global heating and biodiversity loss forces wildlife biologists to focus on anticipating and mitigating impacts upon the natural world. In view of the northward and altitudinal range shifts projected for many Guloninae (Lawler & Hepinstall-Cymerman, 2010) and the associated risks of range fragmentation and reduced population viability, mitigation should prioritize the maintenance and enhancement of habitat connectivity. Efforts should focus on establishing viable ecological networks (Lawton et al., 2010), ideally organized to facilitate anticipated directional and elevational range shifts. This approach is most urgently required in those forested countries that currently experience high rates of resource extraction and continuing primary forest loss (Fisher et al., 2022; Global Forest Review, 2024b); as well as in historically deforested countries where Guloninae populations are constrained by poor availability and connectivity of woodland habitat (e.g. MacPherson & Wright, 2021).
Opportunities to expand and connect Guloninae habitat to maximize population resilience are provided by the global target to halt forest loss and restore 350 million hectares of lost and degraded forests by 2030 (Global Forest Review, 2024c). This will require strategic, landscape-scale approaches in line with the ‘Bigger, Better, More and Joined’ Lawton principles of nature conservation (Lawton et al., 2010). Effective delivery will likely depend upon government-driven land use strategies designed to resolve conflicts between different land use interests striving to achieve climate change adaptation whilst meeting the needs of human society (e.g. Forestry Commission, 2022). A likely hindrance affecting large-scale decarbonization tree-planting is provided by the substantial uncertainties surrounding future climate and economic conditions (Cho et al., 2025).
When making the case for designing reforestation plans to benefit Guloninae, arguments should draw upon evidence that biodiverse forest ecosystems are more resilient to the impacts of the CEE (Thompson et al., 2009) and that Guloninae can deliver ecosystem services, including landscape-scale biological control of invasive species (e.g. Twining et al., 2022). Such benefits should be recognized as part of the expanding field of nature-based solutions to the societal challenges of climate breakdown and biodiversity loss (Seddon et al., 2021).
In rising to the challenges presented by the CEE, conservation biologists should recognize the role that tipping elements may play in the earth’s climate system, notably where these create uncertainty in projections of environmental change. For example, the Atlantic meridional overturning circulation (AMOC) is one such tipping element that, if it collapses under pressure from oceanic freshwater forcing, would trigger a reversal in the current warming of the northern hemisphere, with the greatest cooling apparent in western Europe (van Westen et al., 2024).
Conclusions
The increasingly catastrophic impacts of the Climate and Ecological Emergency will likely define the work of wildlife biologists through the remainder of this century and beyond. So, efforts must focus on maximizing Guloninae resilience by addressing the following tasks at appropriate geographical scales:
- Understanding the drivers of harm to Guloninae
- Predicting and monitoring Guloninae responses to the CEE
- Reporting impacts upon Guloninae to wider society
- Designing and enacting mitigation for Guloninae
- Securing ways of continuing research and conservation work
- Prioritising research and conservation work on the most vulnerable and/or neglected species
These actions may provide a foundation for conservation action which, if enacted effectively by wildlife managers, could mitigate the impacts of the CEE upon Guloninae as they increase in severity through the 21st century (Prato, 2009).
In many countries, the future survival of Guloninae will depend upon the forestry industry and its efforts to adapt to the CEE and, thereby, to meet the needs of human society. Those efforts will involve substantial changes in forestry practice – many of which will directly or indirectly impact Guloninae – as adaptive measures are deployed to combat new or elevated risks to forests of windthrow, wildfires, drought, heat stress, flooding, pests and diseases (e.g. Atkinson et al., 2022; Tew et al., 2024). Nevertheless, in an era of likely great reductions in funding for wildlife research and conservation, climate change adaptation commitments to deliver landscape-scale reforestation provide the best opportunities to mitigate CEE impacts upon Guloninae.
Acknowledgments
We are grateful to members of the Martes Working Group for valuable discussions around Guloninae and the CEE; notably, we thank Jo Burgar for pre-publication permission to reproduce the Guloninae global range map. We thank three anonymous reviewers for their significant contributions to improving this paper. We thank Stacy Coty for help with access to literature and Bryn Evans for editorial input. JB acknowledges the special contribution – by deepening his understanding of the climate and biodiversity crises – of scientists and fellow activists at Extinction Rebellion and journalists at The Guardian newspaper.
Author Contributions
Johnny Birks: conceptualization, original draft, formal analysis, writing, review & editing;
Stephanie Johnstone: writing, review & editing, data curation, formal analysis;
Ed Snell: writing, review & editing, data curation, formal analysis;
Jenny MacPherson: conceptualization, formal analysis, writing, review & editing.
Data Availability
This manuscript did not generate new data and collated data are presented in Tables 1 and 2.
Transparent Peer Review
Results from the Transparent Peer Review can be found here.
Recommended Citation
Birks, J., Johnstone, S., Snell, E., & MacPherson, J. (2025). What is the future for The Martes Complex (Guloninae) in the face of climate change and ecological breakdown? Stacks Journal: 25006. https://doi.org/10.60102/stacks-25006
References
Abramov, A.V. (2016). Gulo gulo. The IUCN Red List of Threatened Species 2016: e.T9561A45198537. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T9561A45198537.en. Accessed on 08 January 2024
Abramov, A.V., Kranz, A., Herrero, J., Choudhury, A. & Maran, T. (2016). Martes foina. The IUCN Red List of Threatened Species 2016: e.T29672A45202514. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T29672A45202514.en. Accessed on 08 January 2024.
Abramov, A.V., Kaneko, Y. & Masuda, R. (2015). Martes melampus. The IUCN Red List of Threatened Species 2015: e.T41650A45213228. https://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T41650A45213228.en. Accessed on 08 January 2024.
Almond, R.E.A., Grooten M. & Petersen, T. (Eds). (2020). Living Planet Report 2020 – Bending the curve of biodiversity loss. WWF, Gland, Switzerland.
Atkinson, G., Morison, J., & Nicoll, B. (2022). Adapting forest and woodland management to the changing climate. UK Forestry Standard Practice Guide. Forest Research, Edinburgh, UK.
Balakrishnan, P. (2005). Recent sightings and habitat characteristics of the endemic Nilgiri Marten Martes gwatkinsii in Western Ghats, India. Small Carnivore Conservation, 33, pp.14-16.
Barnes, M. L., Zhang, Q., Robeson, S.M., Young, L., Burakowski, E. A., Oishi, A. C., Stoy, P. C., Katul, G. & Novick, K. A. (2024). A century of reforestation reduced anthropogenic warming in the Eastern United States. Earth’s Future, 12, e2023EF003663. https://doi.org/10.1029/2023EF003663
Beck, H. E., Zimmermann, N. E., McVicar, T. R., Vergopolan, N., Berg, A., & Wood, E. F. (2018). Present and future Köppen-Geiger climate classification maps at 1-km resolution. Scientific Data 5, 180214. https://doi.org/10.1038/sdata.2018.214
Burgar, J. et al (In Press). A path towards the conservation and recovery of Guloninae species worldwide. Stacks Journal.
Buskirk, S.W. & Powell, R.A. (1994). Habitat ecology of American martens and fishers. Martens, sables and fishers: biology and conservation, pp.297-315.
Buskirk, S.W., Yiqing, M., Li, X. and Zhaowen, J. (1996). Winter habitat ecology of sables (Martes zibellina) in relation to forest management in China. Ecological Applications, 6(1), pp.318-325.
Carr, S. M., & Hicks, S. A. (1997). Are there two species of marten in North America? Genetic and evolutionary relationships within Martes. Martes: taxonomy, ecology, techniques, and management, 15-28.
Carroll, C. (2007). Interacting effects of climate change, landscape conversion, and harvest on carnivore populations at the range margin: marten and lynx in the northern Appalachians. Conservation Biology 21:1092-1104.
Evans, Margaret, Cory Merow, Sydne Record, Sean McMahon, and Brian Enquist. 2016. “Towards process-based range modeling of many species.” Trends in Ecology and Evolution 31 no. 11: 860-871.
Carroll, K.A., Hansen, A.J., Inman, R.M. and Lawrence, R.L. (2021). Evaluating the importance of wolverine habitat predictors using a machine learning method. Journal of Mammalogy, 102(6), pp.1466-1472.
Ceballos, G. and Ehrlich, P. R. (2023). Mutilation of the tree of life via mass extinction of animal genera. Proceedings of the National Academy of Sciences. 120. 39:e2306987120
Cho, F.H.T., Aglonucci, P., Bateman, I.J. and Day, B.H. Resilient tree-planting strategies for carbon dioxide removal under compounding climate and economic uncertainties. Proceedings of the National Academy of Sciences, 122 (10) e2320961122.
Chutipong, W., Duckworth, J.W., Timmins, R.J., Choudhury, A., Abramov, A.V., Roberton, S., Long, B., Rahman, H., Hearn, A., Dinets, V. & Willcox, D.H.A. (2016). Martes flavigula. The IUCN Red List of Threatened Species 2016: e.T41649A45212973. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41649A45212973.en. Accessed on 08 January 2024.
Climate Action Tracker (2023). Press release 5/12/2023: Projected warming almost unchanged for two years as governments push false solutions over climate action. https://climateactiontracker.org/press/release-projected-warming-almost-unchanged-for-two-years-as-governments-push-false-solutions-over-climate-action/ Accessed 31 January 2024.
Copernicus Climate Change Service (2025). Global Climate Highlights 2024. https://climate.copernicus.eu/global-climate-highlights-2024#:~:text=In%202024%2C%2011%20months%20and,C%20and%201.54%C2%B0C. Accessed 30 January 2025.
Cuarón, A.D., Reid, F., Helgen, K. & González-Maya, J.F. (2016). Eira barbara. The IUCN Red List of Threatened Species 2016: e.T41644A45212151. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41644A45212151.en. Accessed on 08 January 2024.
Dawson, N. G., Cook, J. A., Aubry, K. B., Zielinski, W. J., Raphael, M. G., & Proulx, G. (2012). Behind the genes. Biology and conservation of martens, sables, and fishers: a new synthesis, 23-38.
Elsen, P.R. & Tingley, M.W. (2015). Global mountain topography and the fate of montane species under climate change. Nature Climate Change, 5(8), pp.772-776.
Fisher, J.T., Murray, S., Barrueto, M., Carroll, K., Clevenger, A.P., Hausleitner, D., Harrower, W., Heim, N., Heinemeyer, K., Jacob, A.L. and Jung, T.S. (2022). Wolverines (Gulo gulo) in a changing landscape and warming climate: A decadal synthesis of global conservation ecology research. Global Ecology and Conservation, 34, https://doi.org/10.1016/j.gecco.2022.e02019
Fletcher, C., Ripple, W. J., Newsomec, T., Barnard, P., Beamerf, K., Behla, A. Bowen, J., Cooney, M., Crist, E., Field, C., Hiser, K., Karl, D. M., King, D.A., Mann, M. E., McGregor, D. P., Mora, C., Oreskes, N. and Wilson, M. (2024). Earth at risk: An urgent call to end the age of destruction and forge a just and sustainable future. PNAS Nexus, 2024, 3, 1–20 https://doi.org/10.1093/pnasnexus/pgae106
Foden, W.B., Butchart, S.H., Stuart, S.N., Vié, J.C., Akçakaya, H.R., Angulo, A., DeVantier, L.M., Gutsche, A., Turak, E., Cao, L. and Donner, S.D., 2013. Identifying the world’s most climate change vulnerable species: a systematic trait-based assessment of all birds, amphibians and corals. PloS one, 8(6), p.e65427.
Fonda, F., Chiatante, G., Meriggi, A., Mustoni, A., Armanini, M., Mosini, A., Spada, A., Lombardini, M., Righetti, D., Granata, M. and Capelli, E. (2021). Spatial distribution of the pine marten (Martes martes) and stone marten (Martes foina) in the Italian Alps. Mammalian Biology, 101, pp.345-356.
Forestry Commission (2022). Responding to the climate emergency with new trees and woodlands. Forestry Commission https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1057842/WoodlandsTreesForests.pdf Accessed 17th February 2024.
Gardner Charlie J. , Thierry A., Rowlandson W., Steinberger J. K. (2021). From Publications to Public Actions: The Role of Universities in Facilitating Academic Advocacy and Activism in the Climate and Ecological Emergency. Frontiers in Sustainability vol. 2. 10.3389/frsus.2021.679019 ISSN=2673-4524
Geary, W. L., T. S. Doherty, D. G. Nimmo, A. I. T. Tulloch, and E. G. Ritchie (2020). Predator responses to fire: a global systematic review and meta-analysis. Journal of Animal Ecology 89: 955–971.
Global Forest Review (2024a). Forest Loss Indicator. updated April 4, 2024. Washington, DC: World Resources Institute. Available online at https://research.wri.org/gfr/forest-extent-indicators/forest-loss.
Global Forest Review (2024b). Tropical Forest Loss Drops Steeply in Brazil and Colombia, but High Rates Persist Overall. Updated April 4, 2024. Washington, DC: World Resources Institute. Available online at https://research.wri.org/gfr/latest-analysis-deforestation-trends
Global Forest Review (2024c). Deforestation and Restoration Targets Tracker (Beta). updated April 4, 2024. Washington, DC: World Resources Institute. Available online at https://research.wri.org/gfr/forest-targets-tracker
Global Forest Watch (2025). Global Annual Tree Cover Loss. https://globalforestwatch.org/dashboards/global/
Goldenberg, J., Bisschop, K., D’Alba, L. and Shawkey, M.D. (2022), The link between body size, colouration and thermoregulation and their integration into ecogeographical rules: a critical appraisal in light of climate change. Oikos, 2022: e09152. https://doi.org/10.1111/oik.09152
Green, D. S., Martin, M. E., Powell, R. A., McGregor, E. L., Gabriel, M. W., Pilgrim, K. L., Schwartz, M. K. and Matthews, S. M. (2022). Mixed‐severity wildfire and salvage logging affect the populations of a forest‐dependent carnivoran and a competitor. Ecosphere. 13 (1) https://doi.org/10.1002/ecs2.3877
Greenfield, P. (2024). The end of the great northern forests? The tiny tree-killing beetle wreaking havoc on our ancient giants. The Guardian 5th June 2024. https://www.theguardian.com/environment/article/2024/jun/05/collapse-northern-boreal-forests-drought-fire-beetles-climate-crisis-ancient-trees-carbon-sink?CMP=Share_iOSApp_Other
Hargis, C. D., Bissonette, J. A. & Turner, D.L. (1999). The influence of forest fragmentation and landscape pattern on American martens. Journal of Applied Ecology 36:157–72
Helgen, K. & Reid, F. (2016). Martes americana. The IUCN Red List of Threatened Species 2016: e.T41648A45212861. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41648A45212861.en. Accessed on 08 January 2024.
Helgen, K. & Reid, F. (2018). Martes pennanti (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2018: e.T41651A125236220. https://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T41651A125236220.en. Accessed on 08 January 2024.
Herrero, J., Kranz, A., Skumatov, D., Abramov, A.V., Maran, T. & Monakhov, V.G. (2016). Martes martes. The IUCN Red List of Threatened Species 2016: e.T12848A45199169. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T12848A45199169.en. Accessed on 08 January 2024.
Hiltner, E.J. (2022). Changing snow conditions and shifts in American marten (Martes americana) occurrence in Michigan’s Upper Peninsula. Masters thesis, University of Michigan.
Hon, J., Hearn, A.J., Ross, J., Samejima, H., Augeri, D.M., Mathai, J., Mohamed, A., Boonratana, R., Fredriksson, G., Cheyne, S.M. and Heydon, M. (2016). Predicted distribution of the yellow-throated marten Martes flavigula (Mammalia: Carnivora: Mustelidae) on Borneo. Raffles Bulletin of Zoology.
IPBES (2019). Report of the Plenary of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on the work of its seventh session. Paris, 29th April – 4th May 2019. https://files.ipbes.net/ipbes-web-prod-public-files/ipbes_7_10_add.1_en_1.pdf
IPCC (2023): Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland, 184 pp., doi: 10.59327/IPCC/AR6-9789291691647.
IUCN Red List Technical Working Group (2018). Mapping standards and data quality for the IUCN Red List Categories and Criteria. Version, 1(1.16), p.30.Karniski, N., (2014). The effects of snow on American martens (Martes americana) and fishers (Martes pennanti) in the Adirondack Mountains of New York. State University of New York College of Environmental Science and Forestry.
Jolly, W. M., Cochrane, M. A., Freeborn, P. H., Holden, Z. A., Brown, T. J., Williamson, G. J. (2015). Climate-induced Variations in Global Wildfire Danger from 1979 to 2013. Nature Communications. 6, 7537. doi:10.1038/ncomms8537
Karniski, N. (2014). The effects of snow on American martens (Martes americana) and fishers (Martes pennanti) in the Adirondack Mountains of New York. Masters thesis, State University of New York.
Kirk, T.A. and Zielinski, W.J. (2009) Developing and testing a landscape habitat suitability model for the American marten (Martes americana) in the Cascades mountains of California. Landscape Ecology, 24(6), pp.759-773.
Kordosky, J.R., Gese, E.M., Thompson, C.M., Terletzky, P.A., Neuman-Lee, L.A., Schneiderman, J.D., Purcell, K.L. & French, S.S. (2021a). Landscape of stress: Tree mortality influences physiological stress and survival in a native mesocarnivore. PLoS One, 16 (7), p.e0253604.
Kordosky, J.R., Gese, E.M., Thompson, C.M., Terletzky, P.A., Purcell, K.L. and Schneiderman, J.D. (2021b). Landscape use by fishers (Pekania pennanti): core areas differ in habitat than the entire home range. Canadian Journal of Zoology, 99(4), pp.289-297.
Krohn, W. B., Elowe, K. D. & Boone, R. B. (1995). Relations among fishers, snow, and martens: Development and evaluation of two hypotheses. The Forestry Chronicle 71, 97–105.
Kuntze, C. C., Peery, M. Z., Green, R. E., Purcell, K. L. and Pauli, J. N. (2024). Sex and age mediate the effects of rapid environmental change for a forest carnivore, the Fisher (Pekania pennanti). Journal of Mammalogy 105: 13–25.
LaPoint, S. D. (2013) Movement ecology of fishers (Pekania pennanti) within a semi-urban landscape. PhD dissertation, University of Konstanz, Konstanz, Germany
Lawler, J.J., Shafer, S.L., White, D., Kareiva, P., Maurer, E.P., Blaustein, A.R., & Bartlein, P.J. (2009). Projected climate-induced faunal change in the Western Hemisphere. Ecology 90 3 588-597.
Lawler, J.J. & Hepinstall-Cymerman, J. (2010). Conservation Planning in a Changing Climate: Assessing the Impacts of Potential Range Shifts on a Reserve Network. In: Landscape-scale Conservation Planning. Trombulak, S. & Baldwin, R. (Eds). Springer, Dordrecht. https://doi.org/10.1007/978-90-481-9575-6_15
Lawler, J.J., Safford, H.D., & Girvetz, E.H. (2012). Martens and Fishers in a Changing Climate. Pages 371–397 in Biology and Conservation of Martens, Sables, and Fishers. Aubry, K.B., Zielinski, W.I., Raphael, M.G., Proulx, G., & Buskirk, S.W. (Eds) . Cornell University Press, Ithica and London.
Lawler, J.J., Oakes, L.E., Ettinger, A.K. & Lin, Y. (2024). Climate Change: Anticipating and Adapting to the Impacts on Terrestrial Species. Encyclopedia of Biodiversity (3rd Edition). 6: 642-666 https://doi.org/10.1016/B978-0-12-822562-2.00087-6 Accessed 6th February 2024.
Lawton, J.H., Brotherton, P.N.M., Brown, V.K., Elphick, C., Fitter, A.H., Forshaw, J., Haddow, R.W., Hilborne, S., Leafe, R.N., Mace, G.M., Southgate, M.P., Sutherland, W.J., Tew, T.E., Varley, J., & Wynne, G.R. (2010). Making Space for Nature: a review of England’s wildlife sites and ecological network. Report to Defra.
MacPherson, J. & Wright, P. (2021). Long-term strategic recovery plan for pine martens in Britain. Vincent Wildlife Trust Ledbury, UK.
McCann, N. P., Zollner, P. A. & Gilbert, J. H. (2010). Survival of adult martens in northern Wisconsin. Journal of Wildlife Management 74, 1502–1507.
McKay, D.I.A., Staal, A., Abrams, J.F., Winkelmann, R. Sakschewski, B., Loriani, S., Fetzer, I., Cornell, S.E., Rockström, J., & Lenton, T.M. (2022). Exceeding 1.5°C global warming could trigger multiple climate tipping points. Science 377, 6611 DOI: 10.1126/science.abn7950
McNicol, C.M., Bavin, D., Bearhop, S., Bridges, J., Croose, E., Gill, R., Goodwin, C.E., Lewis, J., MacPherson, J., Padfield, D. and Schofield, H. (2020). Postrelease movement and habitat selection of translocated pine martens Martes martes. Ecology and Evolution, 10(11), pp.5106-5118.
Mergey, M., Kaerle, C. and Helder, R., (2023). Variations in density of pine marten Martes martes populations according to forest fragmentation. Mammal Research, 68(4), pp.507-519.
Merz, J.J., Barnard, P., Rees, W.E., Smith, D., Maroni, M., Rhodes, C.J., Dederer, J.H., Bajaj, N., Joy, M.K., Wiedmann, T. & Sutherland, R. (2023). World scientists’ warning: The behavioural crisis driving ecological overshoot. Science Progress 106:3 https://journals.sagepub.com/doi/epub/10.1177/00368504231201372 Accessed on 16 January 2024.
Miyoshi, K. and Higashi, S., (2005). Home range and habitat use by the sable Martes zibellina brachyura in a Japanese cool-temperate mixed forest. Ecological Research, 20, pp.95-101.
Monakhov, V.G. (2016). Martes zibellina. The IUCN Red List of Threatened Species 2016: e.T41652A45213477. https://dx.doi.org/10.2305/IUCN.UK.2016-1.RLTS.T41652A45213477.en. Accessed on 08 January 2024.
Mudappa, D., Jathana, D., & Raman, T.R.S. (2015). Martes gwatkinsii (errata version published in 2016). The IUCN Red List of Threatened Species 2015: e.T12847A86161239. https://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T12847A45199025.en. Accessed on 08 January 2024.
Otte, P. (2021). Habitat selection of beech martens (Martes foina) in relation to prey availability and habitat characteristics in an agricultural landscape (Doctoral dissertation). University of Groningen
Pacifici, M., Foden, W., Visconti, P., Watson, J.E.M, Butchart, S.H.M., Kovacs, K.M., Scheffers, B.R., Hole, D.G., Martin, T.G., Akçakaya, H.R., Corlett, R.T., Huntley, B., Bickford, D., Carr, J.A., Hoffmann, A.A., Midgley, G.F., Pearce-Kelly, P., Pearson, R.G., Williams, S.E., Willis, S.G., Young, B. & Rondinini C. (2015). Assessing species vulnerability to climate change. Nature Climate Change 5, 215–224 . https://doi.org/10.1038/nclimate2448
Pauli, J.N., Manlick, P.J., Tucker, J.M., Smith, G.B., Jensen, P.G. and Fisher, J.T., (2022). Competitive overlap between martens Martes americana and Martes caurina and fishers Pekania pennanti: a rangewide perspective and synthesis. Mammal Review, 52(3), pp.392-409.
Pearson, R.G., Stanton, J.C., Shoemaker, K.T., Aiello-Lammens, M.E., Ersts, P.J., Horning, N., Fordham, D.A., Raxworthy, C.J., Ryu, H.Y., McNees, J. & Akçakaya, H.R. (2014). Life history and spatial traits predict extinction risk due to climate change. Nature Climate Change, 4(3), pp.217-221.
Peel, M. C., Finlayson, B. L., and McMahon, T. A. (2007). Updated world map of the Köppen-Geiger climate classification, Hydrology and Earth System Sciences, 11, 1633–1644, https://doi.org/10.5194/hess-11-1633-2007.
Persson, J., Ordiz, A., Ladle, A., Andrén, H. and Aronsson, M., (2023). Recolonization following past persecution questions the importance of persistent snow cover as a range limiting factor for wolverines. Global Change Biology, 29(20), pp.5802-5815.
Pigot, A.L., Merow, C., Wilson, A., & Trisos, C.H. (2023). Abrupt expansion of climate change risks for species globally. Nature Ecology and Evolution 7, 1060–1071. https://doi.org/10.1038/s41559-023-02070-4 Accessed on 11 January 2024.
Pilgrim, K.L., Green, R.E., Purcell, K.L., Wilcox, T.M., McGregor, E.L., Gleason, L.E., Wasser, S.K., & Schwartz, M.K. (2023). Shifts in fisher (Pekania pennanti) diet in response to climate-induced tree mortality in California assessed with DNA metabarcoding. Journal for Nature Conservation 73 (2023) 126408.
Prato, T. (2009). Evaluating and managing wildlife impacts of climate change under uncertainty. Ecological Modelling. Vol. 220 (7). 923-930.
Proulx, G., Aubry, K.A., Birks, J., Buskirk, S., Fortin, C., Frost, H., Krohn, W., Mayo, L., Monakhov, V., Payer, D., Saeki, M., Santos-Reis, M., Weir, R., & Zielinski, W. (2004). World distribution and status of the genus Martes in 2000. Pages 21-76 in Martens and Fishers (Martes) in human-altered landscapes: an international perspective. D.J. Harrison, A.K. Fuller and G. Proulx (Eds.). Springer Science+Business Media, New York.
Proulx, G. & Aubry, K.B. (2017). The Martes Complex: A monophyletic clade that shares many life-history traits and conservation challenges. In The Martes Complex in the 21st Century: Ecology and Conservation. Zalewski, A., Wierzbowska, I.A., Aubry, K.B., Birks, J.D.S., O’Mahony, D.T. and Proulx, G. (Eds.) Mammal Research Institute, Polish Academy of Sciences, Białowieża.
Pureswaran, D.S., Roques, A. and Battisti, A., (2018). Forest insects and climate change. Current Forestry Reports, 4, pp.35-50.
Rantanen, M., Karpechko, A.Y., Lipponen, A., Nordling, K., Hyvärinen, O., Ruosteenoja, K., Vihma, T., & Laaksonen, A. (2022). The Arctic has warmed nearly four times faster than the globe since 1979. Commun Earth Environ 3, 168. https://doi.org/10.1038/s43247-022-00498-3 Accessed on 23 January 2024.
Rapid Transition Alliance (2021). A new roof for rapid transition – building low-carbon, affordable housing for communities with new ownership models. https://rapidtransition.org/stories/a-new-roof-for-rapid-transition-building-low-carbon-affordable-housing-for-communities-with-new-ownership-models/ Accessed 17th February 2024.
Richards, C.E., Lupton, R.C., & Allwood, J.M. (2021). Re-framing the threat of global warming: an empirical causal loop diagram of climate change, food insecurity and societal collapse. Climatic Change 164, 49. https://doi.org/10.1007/s10584-021-02957-w
Saino, N., Ambrosini, R., Rubolini, D., von Hardenberg, J., Provenzale, A., Hu ̈ppop, K., Hu ̈ppop, O., Lehikoinen, A., Lehikoinen, E., Rainio, K., Romano, M., & Sokolov, L. (2011). Climate warming, ecological mismatch at arrival and population decline in migratory birds. Proc. R. Soc. B 278, 835–842.
Schleussner, C.-F., Nauels, A., Klönne, U., & Hare, B. (2022). Understanding the Paris Agreement’s Long-Term Temperature Goal. Climate Analytics. https://climateanalytics.org/comment/understanding-the-paris-agreements-long-term-temperature-goal Accessed on 24 January 2024.
Schweiger, O., Settele, J., Kudrna, O., Klotz, S., & Kühn, I. (2008). Climate change can cause spatial mismatch of trophically interacting species. Ecology (ESA) 89: 12 https://doi.org/10.1890/07-1748.1 Accessed 6th February 2024.
Shameer, T.T., Mungi, N.A., Backer, S.J., Raman, S., Reddy, S.R., Easa, P.S. and Sanil, R., (2023). Distribution and conservation status of the endemic Nilgiri marten (Martes gwatkinsii). Mammalia, 87(4), pp.360-366.
Scott, A.M., Gilbert, J.H. & Pauli, J.N. (2022). Small mammal dynamics in snow-covered forests. Journal of Mammalogy, 103(3), pp.680-692.
Seddon, N., Smith, A., Smith, P., Key, I., Chausson, A., Girardin, C., House, J., Srivastava, S. and Turner, B. (2021), Getting the message right on nature-based solutions to climate change. Glob. Change Biol., 27: 1518-1546. https://doi.org/10.1111/gcb.15513 Accessed 12 April 2024.
Spencer, W.D, H. Rustigian-Romsos, K. Ferschweiler, and D. Bachelet. (2015). Simulating effects of climate and vegetation change on distributions of martens and fishers in the Sierra Nevada, California, using Maxent and MC1. Pp. 135-149 In: D. Bachelet and D. Turner, eds. Global vegetation dynamics: concepts and applications in the MCI model. Geographical Monograph 214, First Edition. John Wiley & Sons.
Spencer, W.D., Sawyer, S.C., Romsos, H.L., Zielinski, W.J., Thompson, C.M., and Britting, S.A. (2016). Southern Sierra Nevada fisher conservation strategy. Version 1.0. Unpublished report produced by Conservation Biology Institute.
Steel, Z.L., Jones, G.M., Collins, B.M., Green, R., Koltunov, A., Purcell, K.L., Sawyer, S.C., Slaton, M.R., Stephens, S.L., Stine, P. and Thompson, C., 2023. Mega‐disturbances cause rapid decline of mature conifer forest habitat in California. Ecological Applications, 33(2), p.e2763.
Suffice, P., Asselin, H., Imbeau, L., Cheveau, M., & Drapeau, P. (2017). More fishers and fewer martens due to cumulative effects of forest management and climate change as evidenced from local knowledge. J Ethnobiology Ethnomedicine 13, 51. https://doi.org/10.1186/s13002-017-0180-9
Tanigawa, K., Makino, Y., Miura, N., Umeki, K. and Hirao, T. (2022). Scale-dependent habitat selection of sympatric mesocarnivore species in a cool temperate forest in eastern Japan. Mammalian Biology, 102(5), pp.1901-1910.
Tatara, M. (1994). Ecology and conservation status of Tsushima martens, Martes melampus tsuensis. In (S. W. Buskirk, A. S. Harestad, M. G. Raphael and R. A. Powell, eds.) Martens, Sables, and Fishers: Biology and Conservation, pp. 72–279. Cornell University Press, Ithaca, New York
Tew, E.R, Ambrose-Oji, B., Beatty, M., Büntgen, U., Butterworth, H., Clover, G., Cook, D., Dauksta, D., Day, W., Deakin, J., Field, A., Gardiner, B., Harrop, P., Healey, J.R., Heaton, R., Hemery, G., Hill, L., Hughes, O., Khaira-Creswell, P.K., Kirby, K., Leitch, A., MacKay, J., McIlhiney, R., Murphy, B., Newton, L., Norris, D., Nugee, R., Parker, J., Petrokofsky, G., Prosser, A., Quine, C., Randhawa, G., Reid, C., Richardson, M., Ridley-Ellis, D.J., Riley, R., Roberts, J.E., Schaible, R., Simpson, L.E., Spake, R., Tubby, I., Urquhart, J., Wallace-Stephens, F., Wilson, J.D., Sutherland, W.J. (2023). A horizon scan of issues affecting UK forest management within 50 years, Forestry: An International Journal of Forest Research, Volume 97, Issue 3, July 2024, Pages 349–362, https://doi.org/10.1093/forestry/cpad047
Thompson, I., Mackey, B., McNulty, S., & Mosseler, A. (2009). Forest Resilience, Biodiversity, and Climate Change. A synthesis of the biodiversity/resilience/stability relationship in forest ecosystems. Secretariat of the Convention on Biological Diversity, Montreal. Technical Series no. 43, 67 pages.
Thurman, L.L., Stein, B.A., Beever, E.A., Foden, W., Geange, S.R., Green, N., Gross, J.E., Lawrence, D.J., LeDee, O., Olden, J.D. and Thompson, L.M. (2020). Persist in place or shift in space? Evaluating the adaptive capacity of species to climate change. Frontiers in Ecology and the Environment, 18(9), pp.520-528.
Twining, J.P., Lawton, C., White, A., Sheehy, E., Hobson, K., Montgomery, W.I., & Lambin, X. (2022). Restoring vertebrate predator populations can provide landscape-scale biological control of established invasive vertebrates: Insights from pine marten recovery in Europe. Global Change Biology, 28, 5368–5384. https://doi.org/10.1111/gcb.16236
Twining, J.P., Springer, V.L., Cooch, E.G. and Fuller, A.K. (2023). Landscape-scale drivers of tayra abundance in the Ecuadorian Andes. Biodiversity and Conservation, 32(8), pp.2925-2942.
UN Climate Change Conference UK 2021. (2021). Glasgow Leaders’ Declaration on Forests and Land Use. https://webarchive.nationalarchives.gov.uk/ukgwa/20230418175226/https://ukcop26.org/glasgow-leaders-declaration-on-forests-and-land-use/ Accessed on 25 January 2024.
van Westen, R.M., Kliphuis, M., & Dijkstra, H.A. (2024). Physics-based early warning signal shows that AMOC is on tipping course. Science Advances 10, 6 https://www.science.org/doi/10.1126/sciadv.adk1189
Vergara, M., Cushman, S.A., Urra, F., & Ruiz-González, A. (2016). Shaken but not stirred: multiscale habitat suitability modeling of sympatric marten species (Martes martes and Martes foina) in the northern Iberian Peninsula. Landscape Ecol 31, 1241–1260. https://doi.org/10.1007/s10980-015-0307-0
Volkmann, L.A., Hutchen, J. and Hodges, K.E. (2020). Trends in carnivore and ungulate fire ecology research in North American conifer forests. Forest Ecology and Management, 458, p.117691.
Volkmann, L. A. & Hodges, K. E. (2021). Post-fire movements of Pacific marten (Martes caurina) depend on the severity of landscape change. Movement Ecology 9: 49. https://doi.org/10.1186/s40462-021-00286-2
Volkmann, L.A. & Hodges, K.E. (2022). Residual forest structure influences behavior of Pacific marten (Martes caurina) on post-fire landscapes. International Journal of Wildland Fire, 31(4), pp.329-349.
Weeks, B.C., Willard, D.E., Zimova, M., Ellis, A.A., Witynski, M.L., Hennen, M. and Winger, B.M. (2020), Shared morphological consequences of global warming in North American migratory birds. Ecology Letters, 23: 316-325. https://doi.org/10.1111/ele.13434
Wereszczuk, A., Hofmeester, T.R., Csanády, A., Dumić, T., Elmeros, M., Lanszki, J., Madsen, A.B., Müskens, G., Papakosta, M.A., Popiołek, M., Santos‑Reis, M., Zuberogoitia, I., & Zalewski, A. (2021). Different increase rate in body mass of two marten species due to climate warming potentially reinforces interspecific competition. Scientific Reports 11:24164.
Wereszczuk, A., Fedotova A., Marciszak, A., Popiołek, M., Zharova, A. & Zalewski, A. (2023). Various responses of pine marten morphology and demography to temporal climate changes and primary productivity. Journal of Zoology. 319 42-53.
Wright, P.G.R., Croose, E., & MacPherson, J.L. (2022). A global review of the conservation threats and status of mustelids. Mammal Review 52, 3 410-424.
Zalasiewicz, C., Williams, M., Steffen, W., & Crutzen, P. (2010). The New World of the Anthropocene. Environmental Science & Technology 44(7):2228-2231 https://pubs.acs.org/doi/10.1021/es903118j Accessed on 10th April 2024.
Zalasiewicz, J., Waters, C. and Williams, M., (2020). The anthropocene. In Geologic time scale 2020 (pp. 1257-1280). Elsevier.
Zielinski, W. J., Tucker, J. M., & Rennie, K. M. (2017). Niche overlap of competing carnivores across climatic gradients and the conservation implications of climate change at geographic range margins. Biological Conservation, 209, 533-545.
Accepted by 2 of 2 reviewers
Open Access
Peer-Reviewed
Creative Commons
Submitted: 12 April 2024
Accepted: 17 April 2025
Published: 13 June 2025
Funding Information: No funding was received for this study.
Conflicts of Interest: The authors declare no conflicts of interest.



