

Space use and fate of a three-legged coyote – a case study
JH: https://orcid.org/0000-0002-2602-0400
JM: https://orcid.org/0009-0006-8411-9376
2 San Martin Veterinary Services, LLC, Lake Charles, Louisiana, USA
3 College of Forest Resources and Environmental Science, Michigan, USA. https://orcid.org/0000-0002-5104-3719
4 The Canid Project, Baton Rouge, Louisiana, USA. https://orcid.org/0000-0001-6755-0160
Hinton, J.W., San Martin, K., Brzeski, K.E., Murphy, J.J., & Shutt, A.C. (2024). Space use and fate of a three-legged coyote – a case study.The Stacks: 24002. https://doi.org/10.60102/stacks-24002

Abstract photo. Dr. Kelsey San Martin conducting the amputation surgery on LA25M. LA25M suffered a severe injury to his right front foot when captured by biologists. Photo by Amy Shutt.
Abstract
Wildlife rehabilitation is a widespread practice, but it is rarely provided for research animals in wild settings when injuries such as bone fractures occur during field work. Integrating rehabilitation and post-release monitoring with field research involving radio telemetry could improve our ability to rehabilitate wild animals by assessing the efficacy of clinical and rehabilitation techniques. While conducting a study in coastal southwestern Louisiana during 2021–2023 designed to assess coyote (Canis latrans) populations for red wolf (Canis rufus) ancestry, we severely injured a coyote in a foothold trap. Instead of humanely euthanizing the animal, we opted to provide clinical treatment which involved amputating the coyote’s injured forelimb. The three-legged coyote was released with a Global Positioning System (GPS) collar and monitored until his death. Using time local convex hulls and resource selection functions, we observed the three-legged coyote exhibiting similar movement speed and space use as did his three uninjured neighboring GPS-collared coyotes (control animals). However, the amputee coyote exhibited stronger selection for roads and open wetlands than did the control animals, indicating that the amputation may have altered his ability to traverse some land cover types such as areas with increasing vegetation cover. Although the control animals were killed by humans while attempting to enter a fenced-in game preserve, the three-legged coyote avoided entering the same preserve and was presumably killed by an American Alligator (Alligator mississippiensis) within his territory indicating that he avoided areas with high potential for human-coyote conflict. Despite the small sample size of one clinically treated animal, we overcame other common limitations to post-release monitoring studies such as a lack of detailed space use data or use of control animals by using GPS technology on a treated coyote and its neighboring coyotes. Wildlife rehabilitation can provide second chances to animals severely injured by research activities, and we suggest that clinical treatment and rehabilitation should be considered in study designs as rehabilitated animals can maintain good general health and welfare following releases and contribute to local population persistence.
Keywords: amputation, coyote, Canis latrans, foothold trap, movement, rehabilitation, space use
Introduction
For wild carnivores, traumatic injuries such as bone fractures and amputations occur when they are trapped for research, recreation, or population control (Onderka et al. 1990, Rio-Maior et al. 2016, Lamb et al. 2022, Nájera et al. 2024). Although carnivores that suffer traumatic leg injuries may be rehabilitated and released back into the wild, releases are not common and, when they do occur, success is often unknown for several reasons. First, carnivores injured when trapped for purposes of recreation or population control are killed regardless of the general health and welfare of the trapped animal. Second, researchers may opt to humanely euthanize injured carnivores so that injured animals do not experience long-term suffering or painful deaths (Vogelnest 2008, Sikes and Animal Care and Use Committee of the American Society of Mammalogists 2016, Hernandez et al. 2019, Soulsbury et al. 2020). Additionally, disabilities acquired from injuries may negatively impact the long-term fitness and survival of individuals and cause rehabilitated carnivores to become more involved in human-wildlife conflicts (Brown & Tribe 2001, Hernandez 2019). Finally, when carnivores are rehabilitated, post-monitoring studies are limited by small sample sizes and/or lack of institutional finances or experience necessary for conducting research (Hernandez 2019, Cope et al. 2022).
A recent systematic review of the survival of wildlife following rehabilitation indicated that survival during rehabilitation and post-release was species- and context-specific (Cope et al. 2022). These findings are not surprising given that some institutions exist for the purpose of rehabilitating stranded or injured marine mammals, sea turtles, and shore birds and have participated in some post-release studies investigating the efficacy of rehabilitation procedures (Adimey et al. 2016, Wilhelm et al. 2013, Mestre et al. 2014, Baker et al. 2015, Raine et al. 2020, Willette et al. 2023). Despite financial, expertise, and other limitations, post-release evaluation of terrestrial animals following rehabilitation is a field of research needed for providing information on clinical techniques and outcomes of the health and fate of rehabilitated animals (Rio-Maior et al. 2016, Jeong et al. 2021, Nájera et al. 2024). For example, Nájera et al. (2024) reported that forelimb and hindlimb amputations did not affect the movement, foraging, and reproductive behaviors of four free-living amputee felids and suggested that their findings justified “second chances” in the wild for rehabilitated animals. Similarly, Rio-Maior et al. (2016) reported the post-release movements of two rehabilitated Iberian gray wolves (Canis lupus) that suffered severe leg injuries and suggested that wolves can survive in the wild following the rehabilitation of traumatic injuries including limb amputations.
Coyotes (Canis latrans), a moderately-sized North American canid that significantly expanded its geographic range during the 20th century (Hody & Kays 2018, Hinton et al. 2019), have become a commonly rehabilitated species because of their presence in urban areas (Gerht et al. 2011, Hernandez 2019, Van Patter 2022). Recently, the results of an unpublished study involving post-release monitoring of rehabilitated coyotes that survived being hit by cars in Arizona were briefly summarized by Hernandez (2019) and mentioned anecdotally in Lord and Miller (2019). Following surgeries and rehabilitation for long-bone fractures, coyotes were fitted with radio collars and monitored for 12 months post-release. Hernandez (2019) noted that the coyotes were observed scavenging for food in high-density urban areas and were at high risk of suffering similar fates despite being released 32.2 km outside a large city. In Ontario, Canada, Global Positioning System (GPS) radio collars were used to monitor 11 rehabilitated coyotes following their releases (Van Patter 2022). Using a socio-ecological and story-telling framework, Van Patter (2022) detailed post-release monitoring for one of the rehabilitated coyotes. Following an 80-day treatment for sarcoptic mange (Sarcoptes scabiei) and a surgery to remedy a split eyelid and ingrown eyelashes, approximately 10 months of post-release monitoring indicated that the female coyote found her mate and bred successfully.
In Apr 2021, we severely injured a coyote while trapping and radio collaring animals for a regional study along coastal Louisiana designed to assess coyote populations for red wolf (Canis rufus) ancestry (see vonHoldt et al. 2022). Our research team has extensive experience trapping and monitoring coyotes (e.g., Hinton et al. 2015, Ward et al. 2018, Hinton and Chamberlain 2022, Webster et al. 2022, vonHoldt et al. 2022) and some radio-marked coyotes in our previous projects survived forelimb amputations caused by traumatic injuries presumably anthropogenic in origin (e.g., vehicle collision, gunshot, or trap) during their monitoring period with good health and appeared to have contributed to local population persistence. Given that we observed radio-marked coyotes surviving traumatic injuries with some three-legged individuals appearing to us as having lived similar lives as those that did not experience traumatic injuries, we opted to amputate the animal’s leg to preserve its life and released it back into the wild fitted with a GPS radio collar so that we could monitor its activities and potentially know its fate. Additionally, coyotes in this region harbor novel red wolf genetics lost to the extant red wolf population (Heppenheimer et al. 2018, vonHoldt et al. 2022). Therefore, we believed preserving this animal’s life minimized the negative impacts that our research activities may have had on red wolf ancestry in that area while allowing for the coyote to provide us with important insights into the free-ranging behaviors of amputees (Rio-Maior et al. 2016).
Three-leg coyotes are rare and it is believed that three-legged animals will struggle for survival as a result of the inability to catch prey or escape predation (Lord and Miller 2019). Additionally, the rarity of observations and descriptions of three-legged coyotes in the scientific literature may cause us to underestimate the resilience of amputees in the wild. For example, a 1939 study on coyote food habits reported that 1.9% of 8,424 coyotes killed by the United States Biological Survey were three-leg animals who exhibited similar diets as normal coyotes (Sperry 1939). To our knowledge, Sperry (1939) is the only study that explicitly assessed the behavior of three-leg coyotes. More recent research on coyotes reported anecdotal information. Gehrt (2004) noted that a radio-monitored three-legged coyote in Chicago was a transient who traversed nearly 140 km2 and may have been responsible for a number of coyote-human conflicts in the area. When assessing the safety and performance of foothold traps, Onderka et al. (1990) reported that one coyote suffered an amputated limb injury during their study when the coyote escaped with a trap on its foot. After collection, the authors found the coyote to be in good condition with a stomach full of voles despite suffering a debilitating limb injury.
We used GPS telemetry data from a three-legged coyote and three uninjured neighboring coyotes to assess if the three-legged coyote exhibited similar movement rate, space use, and habitat selection as did its uninjured neighbors. Specifically, we provided a descriptive comparison on one amputee coyote and three control coyotes while also conducting one resource selection function analysis to assess differences in selection behaviors between the three control animals and the one amputee. This permitted us to compare the behavior and fate of an amputee with appropriate controls (i.e., neighboring individuals from the wild) to assess the value of rehabilitating research casualties (Hernandez 2019, Cope et al. 2022, Nájera et al. 2024). Given the loss of a forelimb, we expected the three-legged coyote to be displaced from the capture site leading to wide-ranging space use (transiency; e.g., see Rio-Maior et al. 2016). We also expected the three-legged coyote to exhibit reduced movement rates and preference for land cover associated with increased ease of movement compared to the control animals, due to the physical impairment caused by the amputation. Finally, and although not testable due to our small sample size, we assumed the three-legged coyote would more likely die at the hands of humans than the neighboring control animals because slower and wider-ranging movements, increased use of roads, and the potential need to use anthropogenic food sources would expose the three-legged coyote to greater risk of human-caused mortality.
Methods and Materials
Study area
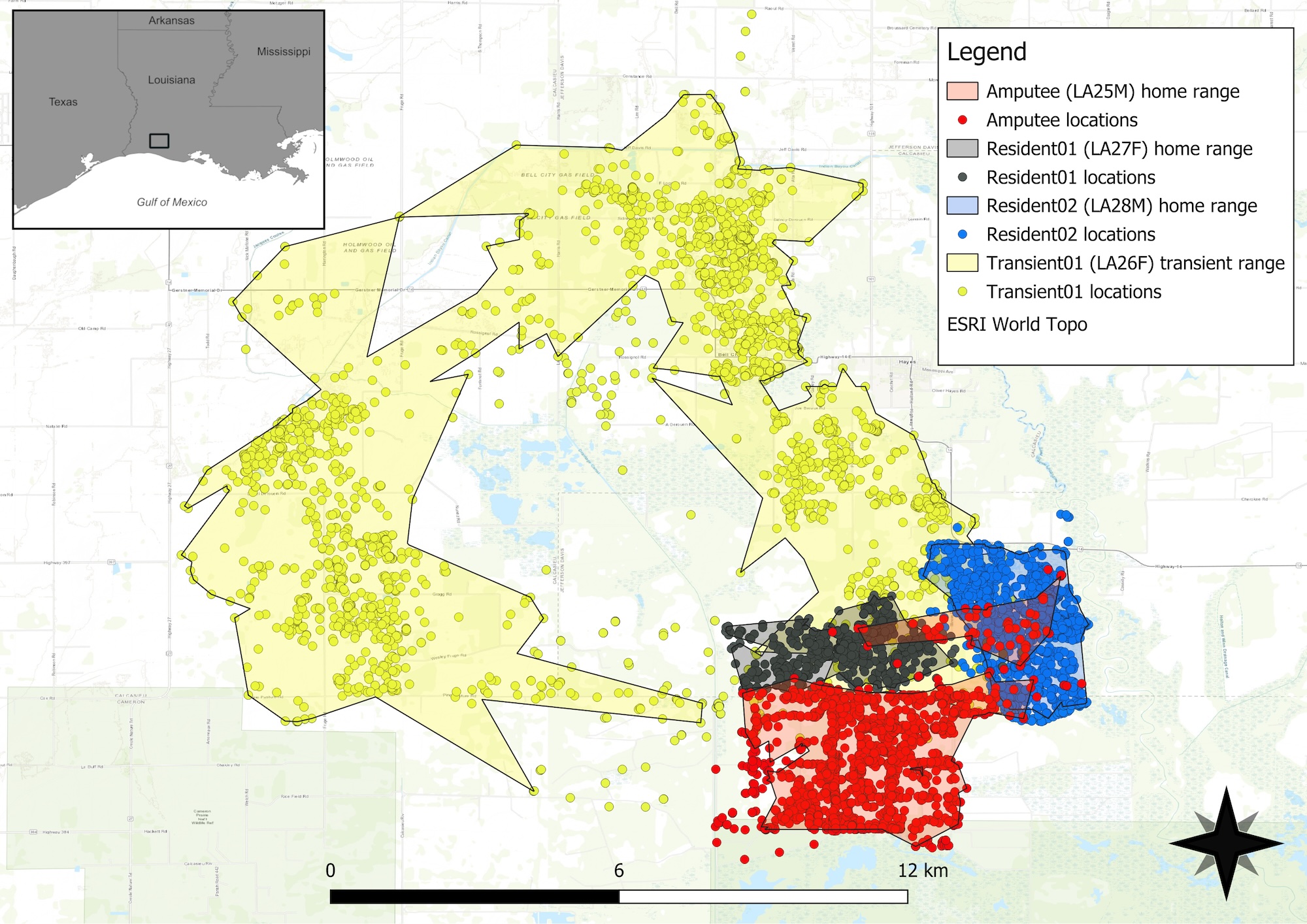
During Apr 2021, we captured four coyotes on 50 km2 of private land (hereafter “the property”) in Jefferson Davis Parish immediately north of Lacassine National Wildlife Refuge (NWR), as part of a larger study of coyotes along the coastal region of southwestern Louisiana (Figure 1; see vonHoldt et al. 2022). Lacassine NWR consisted mostly of natural, freshwater marsh and open water and, to a lesser extent, managed croplands for waterfowl, flooded gum and cypress forest, and tallgrass prairie. The dominant land cover on private lands adjacent to Lacassine NWR was agriculture in the form of rice prairies, crawfish ponds, and domestic cattle operations. The climate at the time of capture was humid subtropical and the area we trapped had mean monthly temperature and precipitation for Apr 2021 of 18.9°C and 12.2 cm, respectively. For more information on the study area, see vonHoldt et al. (2022).
Animal capture and handling
On 27–28 Apr, 2021, we captured four coyotes (LA25M [amputee], LA26F [transient01], LA27F [resident01], and LA28M [resident02]) on the property using foothold traps with offset jaws (Minnesota Brand 550 [MB550], Minnesota Trapline Products, Pennock, MN, USA). We used a combination of anchors and drags when setting foothold traps. Depending on field conditions, foothold traps were anchored into the ground with 40.6 cm super chain stakes or traps were attached to 2.28 kg drags with 2.5 m chains. For example, when brush was available, foothold traps with drags were deployed to allow captured animals to leave the capture site and hide in brush. Although hiding reduced stress for animals, our primary reason for using drags was to avoid having trapped animals remain in the open where they could be more easily seen and killed by humans. When brush was absent, substrate was too difficult to bury drags, or cattle fencing was present, traps were anchored into the ground preventing animals from leaving the capture site.
Our traplines were established along tertiary roads used by agricultural workers and dikes and levees used to divert flood waters from agricultural lands and residential areas. Our capture and handling of animals followed the guidelines approved by the American Society of Mammalogists (Sikes and Animal Care and Use Committee of the American Society of Mammalogists 2016), were approved by the IACUC at Michigan Technological University (#1677987-2), and reported in compliance with the ARRIVE guidelines (Kilkenny et al. 2010).
Except for the injured coyote (LA25M; see Results), captured animals were placed in a Petmate Vari Dog Kennel (Petmate Pet Products, Arlington, TX, USA) using a catchpole and then transported to the maintenance facility on Lacassine NWR to be processed. We processed coyotes at a USFWS facility approximately 5 km away from our trapline to have a secure area away from agricultural activities (e.g., tractor tilling, prescribed burns, and pesticide application) and permit our wildlife veterinarian to thoroughly examine captured animals. Coyotes were removed from kennels using a catchpole, restrained with a muzzle and hobbles, and processed without chemical immobilization. We recorded sex, weight, and body measurements for all animals (Hinton and Chamberlain 2014) and estimated age by tooth wear and body condition (Gier 1968, Gipson et al. 2000). We categorized animals ≥2 years old as adults, 1–2 years old as juveniles, and ≤1-year-old as pups. To assess animals for red wolf ancestry, we collected 5 ml of whole blood from the cephalic veins and stored it in Longmire buffer (vonHoldt et al. 2022). Prior to release at their capture sites, we fitted coyotes with Lotek LiteTrack Iridium 360 GPS collars (Lotek, Newmarket, ON, Canada) scheduled to record a location every 4 hours throughout the year.
Clinical treatment of amputee coyote
The amputee was managed in accordance with the veterinarian hospital’s typical standard care for animals undergoing forelimb amputation. Prior to surgery, he was sedated using Telazol (tiletamine and zolazepam) in combination with medetomidine intramuscularly to provide adequate extended sedation. Upon sedation, the open wound on the dorsal surface of the right front forelimb and over the metacarpal bones was assessed. Metacarpals II through V all sustained complete open fractures at the distal aspects. During this period, we recorded body measurements, sampled blood for genomic analysis, and fitted the injured coyotes with a GPS radio collar.
The forelimb amputation was performed via disarticulation of the scapulohumeral joint without removing the scapula because the residual limb below this level served no useful purpose and would be prone to abrasions and infections if he attempted to use the limb. During post-operation, the amputee received buprenorphine [0.6 mg/ml] intravenously and carprofen [50 mg/ml] subcutaneously for pain management. The amputee also received cefovecin [80 mg/ml] as a prophylactic antibiotic that lasts 10–14 days in most domestic canines. The amputee was allowed to recover in the transport kennel without reversal medication to decrease the chances of a dysphoric recovery and possible self-harm. Given that the amputee was a wild coyote to be re-released into the wild, no post-operative bandaging was provided.
Space use and resource selection
To evaluate whether the amputation affected the space use behaviors of the amputee, we compared his space use metrics to those of the three nearest neighboring coyotes. Specifically, we evaluated the influence of the amputation on the amputee’s space use and habitat selection using 3 metrics: movement rate (km/h), home range size (km2), and coefficients of resource selection. We only included the three coyotes (transient01, resident01, resident02; Table 1) in our analysis because they were neighboring animals with GPS radio collars and two of them (resident01 and resident02) co-occurred on the property with the amputee. Due to our small sample size (one amputee and three healthy control animals), we did a qualitative comparison of metrics among coyotes and only tested for the effect of the amputation on coyote resource selection. We also provided a qualitative narrative of their space use behaviors and fates.
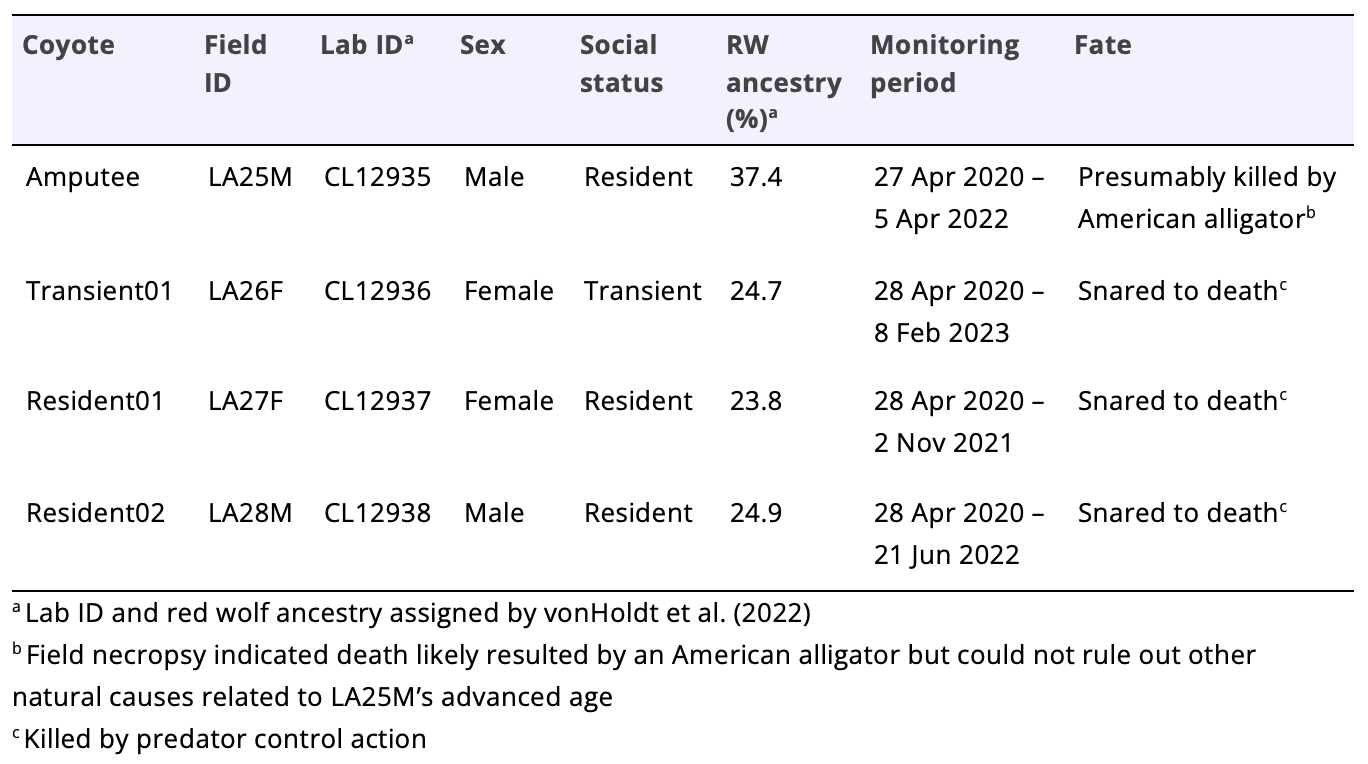
Table 1. Comparison of number of individual participants’ free-listing and pile sorting responses for like/dislike and tolerance/intolerance. All participants (n=69) classified these species based on tolerance, but the sample size for like/dislike varied based on how many participants included the species on their free-lists. Results of Fisher’s exact test are displayed for cases with large enough sample sizes to test for correlation and p-values less than 0.05 are denoted with an asterisk.
We calculated daily distances traversed by coyotes using the “adehabitatLT” library (Calenge 2006) in R 3.6.1 (R Core Team 2019) to create movement trajectories. Movement rates were then estimated as km/h and calculated by converting sequential point locations to line segments and attributed to the midpoint in time between sequential points rounded to the nearest hour.
Using the “T-LoCoH” package (Lyons & Getz 2018) in R, we modeled composite home ranges using time-local convex hulls (T-LoCoH) that advance the traditional LoCoH method, a non-parametric utilization distribution (UD) method that constructs convex hulls associated with each point and its nearest neighbors (Getz and Wilmers 2004; Getz et al. 2007; Lyons et al. 2013). T-LoCoH incorporates a time-scaled distance metric to sort and progressively merge isopleths which addresses spatial and temporal autocorrelation in location data (Schweiger et al. 2015, Lyons et al. 2013). The space (X/Y coordinates) and time (timestamps) components of the time-scaled distance metric are weighted by s, a dimensionless scaling factor of the distance an individual animal could have traveled during the time interval (Lyons & Getz 2018). We selected an s value such that 60% of the polygons were time-selected for each coyote to ensure comparisons between animals were possible (Lyons and Getz 2018). We used the adaptive localized convex hull method (a-method) for identifying nearest neighbors and chose the 50% and 95% aggregations of polygons, or isopleths, to represent the core area and home range, respectively. However, one of our control coyotes was a transient and did not maintain a core area and home range. Unlike resident coyotes that belong to breeding pairs or packs and defend territories, transients are solitary individuals that exhibit both localized and long-distance movements because they are searching for mates and breeding opportunities (Hinton et al. 2015, Morin & Kelly 2017). Consequently, we considered the 50% and 95% isopleths of transient01 as its biding area and transient range, respectively (Hinton et al. 2015).
To develop resource selection functions (RSFs), we followed a 3rd-order resource selection design (Johnson 1980; Manly et al. 2002) to examine the relationship between land cover and coyote space use with their home and transient ranges. We used individual coyotes as sampling units and measured resource availability for each animal. We used Euclidean distance analysis (EDA) to compare estimated animal locations (GPS telemetry locations) and distances from these locations to land cover types of interest with the mean distance to these land cover types across the home and transient ranges (Benson 2013). We used a systematic sampling approach by creating a 30m x 30m raster for each of the land cover types across our study area and calculated the distance from the center of each pixel to the closest representation of that land cover type. Within home and transient ranges, we then used Euclidean distances to measure distances of telemetry locations (use) and systematic random locations (availability) from each land cover type (Benson 2013, Benson et al. 2021).
We overlaid GPS locations onto 30-m resolution digital maps of four land cover classes from national and state databases that likely influenced coyote habitat selection. We used the United States Geological Survey (USGS) 2020 National Land Cover Data set (NLCD) to develop three distance-based land cover classes – distance to open dry cover (classes 52, 71, 81-82), distance to open wetlands (class 95), and distance to woody wetlands (class 90). We acquired a 2020 road layer from the Louisiana Department of Transportation to create a distance to roads land cover class. We created distance raster maps for these four spatial predictors using the Euclidean distance tool in QGIS 3.30 to calculate the distance from every 30-m pixel to the closest landscape feature.
To model resource selection, we used a use-availability framework with generalized linear mixed models (GLMMs) implemented in the R package ‘lme4’ (version 3.4.1) with a binary (0 = available, 1 = used) response variable (Bates et al. 2015). Because we used distance-based variables, we inferred “selection” when known (used) locations were closer to land cover types than were systematic (available) locations as indicated by negative coefficient values (decreasing distances). We constructed several models to test our prediction that LA25M would exhibit greater selection for roads and open land cover types than would the three uninjured coyotes. We considered models with (1) all resource variables and no interactions, (2) interactions between resource variables and a dummy-coded variable from amputation (0 = no amputation, 1 = amputation), and (3) the null model. Interactions with the dummy-coded variable for amputation allowed us to evaluate differences in resource selection between LA25M and the three uninjured coyotes. We calculated the difference in Akaike’s Information Criterion (ΔAIC) between these models to evaluate relative support and made inferences on models with ΔAIC<10 (Bolker et al. 2009). Before modeling, we rescaled land cover variables by subtracting their mean and dividing them by 1 standard deviation.
Results
Capture, trap injury, space use, and fate of the amputee
We captured the amputee on 27 Apr 2021 in the south-central area of his home range (Figure 1). At 0645, we located the amputee lying prone along the wall of a canal with his right front foot caught in a foothold trap. We suspected that the amputee was captured between 0000–0400 27 Apr 2021 following our nightly inspection of traplines in which we found our traps undisturbed between 2200-2330 26 Apr 2021. After the amputee sprung our trap on his foot, he pulled the drag out of the ground and fled to adjacent brush where the drag and chain failed to become entangled in vegetation. Consequently, he then continued pulling the drag for approximately 600m before it became entangled in brush on the edge of a canal.
At the time of the amputee’s capture, the rice field was dry and barren enabling him to pull the drag across an extensive open area. However, because the sheared wedge points and kickers on the drag created resistance by stabbing and turning topsoil, pulling the drag for approximately 600m caused prolonged trauma to the amputee’s captured foot that would otherwise have been minimized if the drag was entangled in the brush immediately adjacent to the trap site. Once the amputee crossed the rice field and climbed up the levee, the drag became entangled in brush along the canal edge where he remained until we located him. The injury to the amputee’s right front leg was a nearly complete amputation with the foot held on by a small piece of tendon (Figure 2). Given the severity of his injury, the amputee was likely in shock and physically exhausted when approached by us and, because of his condition, did not attempt to flee or resist handling when we removed the trap from his leg and placed him in the kennel. The amputee was then transported to the veterinary hospital in Lake Charles, Louisiana, for treatment.
Following his amputation surgery (see Methods), we transported the amputee to the property for release on a relatively small and grassy oil pad with restricted access and adjacent woodlots. Oil pads are areas in which oil and gas production took place on the property and experienced the least amount of human activity. At approximately 2000 27 Apr 2021, we removed the door from the amputee’s kennel and positioned the kennel toward the woodlot and left the area. By positioning the kennel toward the woodlot and allowing the amputee to exit the kennel of his own choice, we believed it increased the likelihood that he would find cover and concealment when leaving. Sometime between 2000-2200 27 Apr 2021, he exited the kennel and loafed in the adjacent woodlot from 2200 27 Apr 2021–0200 29 Apr 2021. This was confirmed when the lead author inspected the kennel and listened for the amputee’s collar while conducting a late-night check of the trapline. Had the amputee remained in the kennel, we would have secured it and taken the amputee back to the veterinary hospital for further evaluation. Between 0200-0600 29 Apr 2021, the amputee left the woodlot and moved back to the core area of his home range. By 4 May 2021, the amputee was traversing his entire home range.

Of the four canids, the amputee had the greatest amount of red wolf ancestry (Table 1) and his size was that of a typical eastern coyote (Table 2). His incisor, canine, and carnassial teeth were worn nearly to the gum line and his overall body condition indicated that he was approximately 10 years old. Given his advanced age, we classified him as a resident animal which was later confirmed via GPS telemetry data.
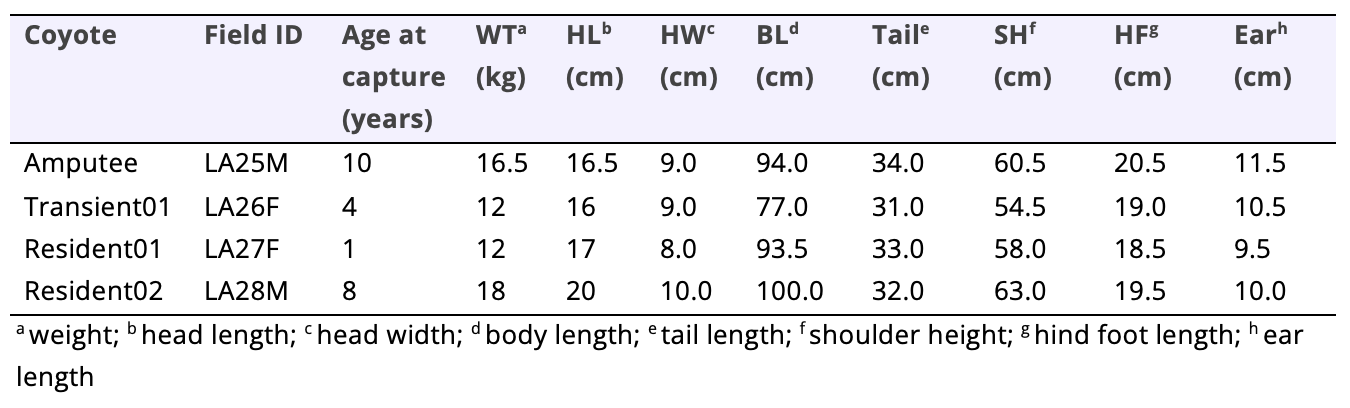
Table 2. Description of four coyotes fitted with Global Positioning Systems (GPS) collars during a study assessing coyote populations along coastal Louisiana for red wolf ancestry (vonHoldt et al. 2022). Each animal was given a unique field and lab ID, sexed (male or female), and social status was assigned following four months of monitoring (Hinton et al. 2015). Location data was collected at 4-hour intervals and telemetry monitoring occurred until study animals died.
Following his release, the amputee exhibited strong fidelity to the area where he was captured. He was monitored for 343 days and maintained a home range and core area of 19.9 km2 and 4.4 km2, respectively. His average rate of movement was 0.14 km/hr (0.13–0.15). He appeared to have been temporarily displaced from the core area of his home range from 27 Nov 2021–3 Feb 2022.
On 14 Apr 2022, we found an American alligator (Alligator mississippiensis) in possession of the amputee’s carcass and telemetry locations indicated that the alligator cached the amputee’s carcass in several different locations prior to us recovering it. Despite observing significant tissue damage to the amputee, the amputation site was one of the few places that remained intact. The surgical incision appeared to have healed well and there was no evidence of sores or new related wounds at the surgical site nor significant muscle atrophy over the right scapula. The lack of evidence of trauma commonly associated with shooting or vehicle collision caused us to believe that the amputee was either killed by an American alligator or scavenged by one following a natural death related to his advanced age. Further leading us to believe the amputee was likely killed by the American alligator in possession of his carcass, it was reported to us that alligators on the property had killed domestic dogs in the past (D. Aucoin, personal communication).
Captures, space use, and fates of the control coyotes
All three control animals (transient01, resident01, and resident02) were captured between 2100–2300 28 Apr 2021. Based on tooth wear and overall body conditions, we estimated the ages of transient01, resident01, and resident02 to be 4-years-old, 1-year-old, and 8-years-old, respectively. Despite significant amounts of red wolf ancestry for the control animals (~24%; Table 1), the two females (transient01 and resident01) were small-framed animals appearing more like western coyotes in size than eastern ones (Table 2, Figure 2). Resident02 was the largest of the four captured coyotes and appeared more like a typical eastern coyote in size (Table 2). Like the amputee, resident02 was estimated to be an old animal due to the significantly worn incisor, canine, and carnassial teeth.
We monitored the three control animals between 188–651 days (Table 1). During her entire monitoring period, transient01 exhibited extensive shifting space use with little fidelity to areas which led us to classify her as a transient (Figure 1). She had a transient range of 108.7 km2 and a biding area of 9.6 km2. Her average rate of movement was 0.17 km/hr (95% CI = 0.16–0.17). Resident01 and resident02 exhibited stable localized movements characteristic of defended space and field evidence (e.g., sightings and tracks) indicated that both animals were with other coyotes which led us to classify them as residents (Figure 1). Resident01 and resident02 had home ranges of 6.7 km2 and 9.3 km2 and core areas of 1.3 km2 and 1.9 km2, respectively. The average rate of movement for resident01 and resident02 were 0.12 km/hr (95% CI = 0.11–0.13) and 0.17 km/hr (95% CI = 0.16–0.17), respectively. All three control animals were snared to death along the property’s game fence located immediately north of resident01’s home range.
Habitat selection by amputee and control coyotes
Model fit improved substantially by accounting for the amputation and fitting interactions between land cover type and amputation to explicitly test for differences in habitat selection between the amputee and the control coyotes, providing support that the amputation affected resource selection (Tables 3 and 4). There was broad selection for open dry cover (β = -0.330, SE = 0.023, p < 0.001) and woody wetlands (β = -0.322, SE = 0.023, p < 0.001) and avoidance of roads (β = 0.051, SE = 0.016, p = 0.002). However, the amputee selected for roads (β = -0.612, SE = 0.067, p < 0.001) and open wetlands (β = -0.399, SE = 0.048, p < 0.001) while avoiding woody wetlands (β = 1.088, SE = 0.052, p < 0.001). In other words, the amputee selected for roads and open wetlands while avoiding woody wetlands, whereas the control animals selected for open dry cover and woody wetlands while avoiding roads.

Table 3. Comparison of model fit among the null model, and models with and without interactions used to test the hypotheses about the effect of an amputation on coyote 3rd-order resource selection in southwestern Louisiana, USA, 2021–2023. Shown are Akaike’s Information Criteria for small sample sizes (AICc), deviance, and differences among AICc (ΔAIC).
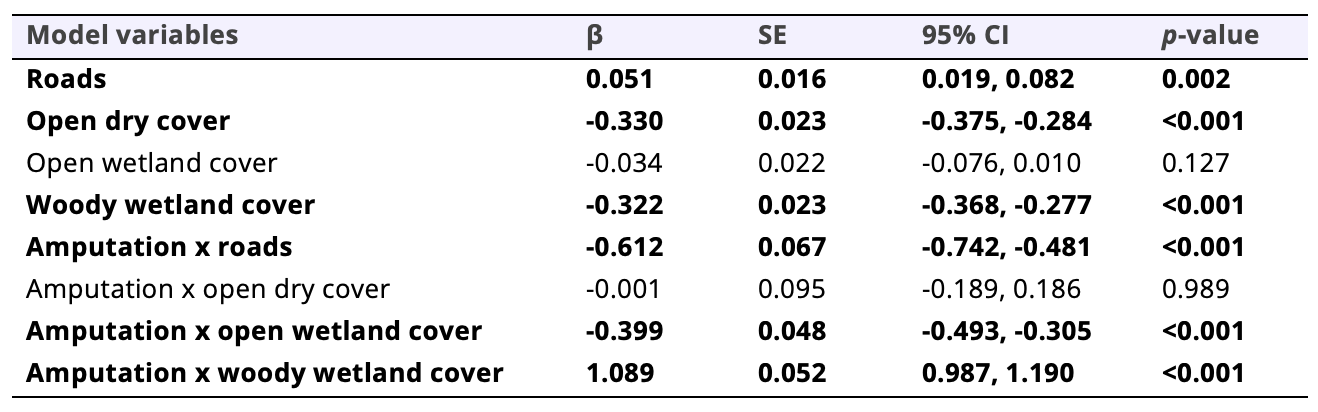
Table 4. Summary of results from generalized linear mixed models with for the full 3rd-order resource selection model with interactions for four coyotes in southwestern Louisiana, USA, during 2021–2023. Shown are β coefficients, standard error (SE), 95% confidence intervals (CI), and p-values. Bolded rows are predictors with statistical support.
Discussion
We successfully amputated the forelimb of a coyote who suffered a severe leg injury when trapped by us and returned it to the wild where it survived for 11 months. We believe that our treatment was effective in ensuring the recovery of the amputee’s health and the loss of his forelimb may not have significantly altered his status as a resident animal. For example, we failed to observe any significant differences in movement rate and space use between the amputee and the control coyotes that could be attributed to the amputation. However, the amputee displayed considerably stronger selection for roads and avoidance of woody wetlands than did the control animals. We interpreted the amputee’s avoidance of woody wetlands as an indicator that the amputation likely influenced his selection for cover types that improved his ease of movement and overall mobility such as those with decreasing vegetation structure. The three control animals were snared to death along the property’s game fence whereas the amputee suffered a non-anthropogenic death presumably by an American alligator. Similar to positive findings reported in other post-release studies of carnivore amputees (Rio-Maior et al. 2016, Jeong et al. 2021, Nájera et al. 2024), we offer that our observations provide evidence that some rehabilitated coyotes with amputations may be able to recover and survive in the wild.
Following his release, the amputee remained under canopy cover for two days before returning to areas proximate to where he was captured, indicating that he was not displaced due to the amputation. Over the course of 11 months, the amputee exhibited space use of a resident animal except between 27 Nov 2021–3 Feb 2022 in which he resided along the western and northern edges of his territory while making several brief excursions to the north into resident01’s and resident02’s home ranges (Figure 1). Given his advanced age, we assumed that he may have lost his breeder position in the pack and was temporarily displaced during the breeding season (January–February) likely returning when the female breeder was pregnant and no longer in estrus (Carlson and Gese 2008). We are uncertain if the amputee was the breeding male before his injury because a larger and presumably younger male was observed routinely traversing his territory when we trapped the area. Nevertheless, the amputee’s absence from the area for most of the breeding season indicates that he was likely a non-breeding resident and may have had familial relationships with the other coyotes that co-occurred with him.
Greater space use by transient01 when compared to the amputee was the result of her biding and exploratory space use associated with being a transient whereas the amputee displayed strong re-visitation and fidelity to an area typical of a resident (Hinton et al. 2015, Morin & Kelly 2017). However, the amputee’s home range was approximately 3x and 2x the size of resident01’s and resident02’s home ranges, respectively, though the amputee’s home-range size was likely inflated because of his temporary displacement from the core area of his pack’s territory for approximately 9 weeks. When we compared space use of the amputee and resident01 for May–Aug 2021, when resident01’s collar was functional, the amputee’s summer home range was nearly 2x larger than hers (12.7 km2 vs 6.7 km2) indicating that his temporary displacement accounted for a 36% increase in his total space use during our monitoring efforts. Because the amputee’s movement rate was comparable to the control animals, we suggest that the amputee compensated adequately for his missing forelimb and likely resumed ranging behaviors typical of coyotes.
Land cover in the amputee’s home range was predominantly rice fields with some early successional/fallow cover and irrigation dikes with coastal bottomland forests encompassing some of the westernmost areas. Levees marked the outermost boundaries of his home range in all cardinal directions. Within this mosaic of land cover, he mostly loafed in early successional/fallow cover surrounded by rice fields during the day and then relied heavily on farm roads to traverse his territory at night. He was periodically seen moving along roads, levees, and open rice fields during early morning hours by the property’s tenet farmers (D. Aucoin, personal communication). Although the amputee displayed similar space use behaviors as did resident01 and resident02 via revisitation to preferred loafing and foraging areas, the amputee displayed considerably stronger selection for roads and avoidance of woody wetlands such as coastal bottomland forests than did resident01 and resident02. We believe the amputation made it difficult for the amputee to traverse areas with uneven ground or vegetation cover or swim across irrigation dikes. Indeed, his selection for roads to navigate around land cover types such as woody wetlands that his neighboring coyotes routinely traversed may have increased his home range size and increased his contact with American alligators that were common in irrigation canals adjacent to some roads in his home range.
All four radio-collared coyotes died during monitoring. Interestingly, the amputee was the only one of the four animals to die from a presumed non-anthropogenic cause and to have died within his home range. The other three coyotes were snared to death along the same game fence on the property. Although transient01 did not have a home range, the locations where resident01 and resident02 were snared along the fence were outside of their home ranges. Given resident01’s young age, it is likely that she may have been snared during an excursion from her territory. We suspect that resident02 may have been drawn to the area when a pack member, such as a pre-dispersing offspring, was snared on the game fence because coyotes were routinely snared to death along the fence (D. Aucoin, personal communication) and resident02 exhibited no excursions from his territory during his nearly 14-month monitoring period. For example, we genetically sampled five coyotes that were snared along the game fence during 23 Feb 2023–24 Mar 2023 and it is unknown if those coyotes belonged to packs inhabiting the property. Nevertheless, during his extraterritorial excursions, the amputee frequented the same areas proximate to the game fence where transient01, resident01, and resident02 were trapped and killed. The amputee’s avoidance of the snares along the game fence and the absence of his telemetry locations in the property’s game preserve indicated that he did not attempt to enter the preserve. This avoidance may be due to more complex behavioral factors related to learning and exploratory behavior vis-á-vis his experience of being trapped and handled by humans (Young et al. 2022, Barrett et al. 2019). Given that the control animals were also trapped and handled by us and that resident02 and the amputee were estimated to be 8 years of age or older and with similar experiences traversing the property and its surrounding areas, we offer that the amputee’s amputation did not affect his ability to avoid areas with high potential for human-coyote conflict.
Little is known about the outcome of the health and fate of rehabilitated wildlife following their release unless individuals are marked and monitored. Recent studies have reported positive outcomes for canids (Rio-Maior et al. 2016), bears (Jeong et al. 2021), and felids (Nájera et al. 2024) that were rehabilitated with leg amputations and released back into the wild. Radio collaring animals is one of the most common and traditional research activities in wildlife studies (Millspaugh and Marzluff 2001, Kays et al. 2015), and with this type of research, injuries caused by trapping and handling occur requiring biologists to make decisions as to whether research casualties are humanely euthanized or rehabilitated and released. Herein, we described the clinical treatment and post-release monitoring of a coyote who suffered an amputation to its forelimb in one of our foothold traps. By doing so, we addressed several limitations in post-release monitoring such as detailed movement, space use, and resource selection, survival and known fate, and a control group of animals. Despite our small sample size, we believe that integrating rehabilitation and post-release monitoring into wildlife research involving the radio collaring of animals can provide information needed to modify and improve clinical techniques by addressing the uncertainties of specific clinical and surgical techniques on the health of rehabilitated animals and the potential impacts of rehabilitated animals on the local receiving population (Jeong et al. 2021). We accomplished integration by justifying the use of wildlife veterinarians to minimize injury or mortality of captured wildlife in our animal use protocols (AUP). Our AUP was required by the Louisiana Department of Wildlife and Fisheries to conduct field research in Louisiana. These institutional approvals allowed us to seek out collaboration with wildlife veterinarians and wildlife rehabilitators during our research. However, we recognize that our approach may not always be wholly applicable to field research, partly because the welfare of the individual animal is, depending on philosophical views of human-animal relations, considered secondary to that of the species as a whole (de Mori 2019, Lynn et al. 2023) and post-release monitoring of long-term effects such as “physical injuries with behavioral and physiological responses” are relatively challenging and often cost-prohibitive (Powell & Proulx 2003).
Acknowledgments
We thank J. Zaunbrecher for providing us access to The Lacassane Company land and support to study coyotes on their properties. We also thank D. Aucoin for updating us on coyote activity on Lacassane Company land and recovering the carcasses and radio collars of our study animals. We also thank the U.S. Fish and Wildlife Service staff at Lacassine National Wildlife Refuge for the use of their facilities and permission to work there. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement. This study was funded by the US Fish and Wildlife Service (F20AC11140-00) and Wolf Conservation Center.
Author Contributions
Joseph W. Hinton: Project administration, field work, conceptualization, investigation, writing – original draft, visualization, and review and editing
Kelsey San Martin: Clinical care, surgery, and review and editing
Kristin E. Brzeski: Project administration, lab work, and review and editing
Jazmin J. Murphy: Original draft and review and editing
Amy Shutt: Field work, visualization, and review and editing
Data Availability
All data from this study are provided here.
Transparent Peer Review
Results from the Transparent Peer Review can be found here.
Recommended Citation
Hinton, J.W., San Martin, K., Brzeski, K.E., Murphy, J.J., & Shutt, A.C. (2024). Space use and fate of a three-legged coyote – a case study. The Stacks: 24002. https://doi.org/10.60102/stacks-24002
References
Adimey, N. M., Ross, M., Hall, M., Reid, J. P., Barlas, M. E., Diagne, L. W. K., & Bonde, R. K. 2016. Twenty-six years of post-release monitoring of Florida manatees (Trichechus manatus latirostris): Evaluation of a cooperative rehabilitation program. Aquatic Mammals 42: 391.
Baker, L., Edwards, W., & Pike, D. A. 2015. Sea turtle rehabilitation success increases with body size and differs among species. Endangered Species Research 29: 13–21.
Bates, D., Mächler, M., Bolker, B., & Walker, S. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67: 1–48.
Barrett, L. P., Stanton, L. A., & Benson-Amram, S. 2019. The cognition of ‘nuisance’ species. Animal Behaviour 147: 167–177.
Benson, J. F. 2013. Improving rigour and efficiency of use‐availability habitat selection analyses with systematic estimation of availability. Methods in Ecology and Evolution 4: 244–251.
Benson, J. F., Abernathy, H. N., Sikich, J. A., & Riley, S. P. D. 2021. Mountain lions reduce movement, increase efficiency during the Covid-19 shutdown. Ecological Solutions and Evidence 2: e12093.
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., & White, J. S. S. 2009. Generalized linear mixed models: a practical guide for ecology and evolution. Trends in Ecology and Evolution 24: 127–135.
Brown, P. R., & Tribe, A. 2001. ‘Well it sounded like a great idea at the time’: wildlife rehabilitation and relocation in Australia. Pages 187–197 in A. Martin, and L. Vogelnest, editors. Veterinary conservation biology: Wildlife health and management in Australasia. Proceedings of a joint conference. Sydney, Australia.
Calenge, C. 2006. The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197: 516–519.
Carlson, D. A., & Gese, E. M. 2008. Reproductive biology of the coyote (Canis latrans) integration of mating behavior, reproductive hormones, and vaginal cytology. Journal of Mammalogy 89: 654–664.
Cope, H. R., McArthur, C., Dickman, C. R., Newsome, T. M., Gray, R., & Herbert, C. A. 2022. A systematic review of factors affecting wildlife survival during rehabilitation and release. PLoS ONE 17: e0265514.
de Mori, B. 2019. Animal testing: The ethical principle of the 3Rs from laboratories to “field” research with wild animals. Ethics & Politics 21: 553–570.
Gehrt, S. D. 2004. Chicago coyotes part I. Wildlife Control Technology 11: 24–26.
Gehrt, S. D., Brown, J. L., & Anchor, C. 2011. Is the urban coyote a misanthropic synanthrope? The case from Chicago. Cities and the Environment 4: 3.
Getz, W. M., & Wilmers, C. C. 2004. A local nearest-neighbor convex-hull construction of home ranges and utilization distributions. Ecography 27: 489–505.
Getz, W. M., Fortmann-Roe, S., Cross, P. C., Lyons, A. J., Ryan, S. J., and Wilmers, C. C. 2007. LoCoH: nonparametric kernel methods for constructing home ranges and utilization distributions. PLoS ONE 2: e207.
Gier, H. T. 1968. Coyotes in Kansas. Kansas State College Agricultural Experimental Station Bulletin 393: 1–118.
Gipson, P., Ballard, W., Nowak, R., and Mech, L. 2000. Accuracy and precision of estimating age of gray wolves by tooth wear. Journal of Wildlife Management 64: 752–758.
Heppenheimer, E., Brzeski, K.E., Wooten, R., Waddell, W., Rutledge, L.Y., Chamberlain, M.J., Stahler, D.R., Hinton, J.W. and VonHoldt, B.M. 2018. Rediscovery of red wolf ghost alleles in a canid population along the American Gulf Coast. Genes 9: 618.
Hernandez, S. M. 2019. Postrehabilitation release monitoring of wildlife. In pages 123–127, Hernandez, S. M., Barron, H. W., Miller, E. A., Aguilar, R. F., and Yabsley, M. J., eds. Medical management of wildlife species: a guide for practitioners. John Wiley & Sons, Hoboken, NJ.
Hernandez, S. M., Barron, H. W., Miller, E. A., Aguilar, R. F., and Yabsley, M. J., eds. 2019. Medical management of wildlife species: a guide for practitioners. John Wiley & Sons, Hoboken, NJ.
Hinton, J. W., and Chamberlain, M. J. 2014. Morphometrics of Canis taxa in eastern North Carolina. Journal of Mammalogy 95: 855–861.
Hinton, J. W., and Chamberlain, M. J. 2022. Evidence of reduced abundance, density, and survival of coyotes under federal management for red wolf recovery. Ecosphere 13: e4216.
Hinton, J. W., van Manen, F. T., and Chamberlain, M. J. 2015. Space use and habitat selection by resident and transient coyotes (Canis latrans). PloS ONE 10: e0132203.
Hinton, J. W., Heppenheimer, E., West, K., Caudill, D., Walch, M., Kilgo, J. C., Mayer, J., vonHoldt, B., and Chamberlain, M. J. 2019. Geographic patterns in morphometrics and genetic variation for coyote populations with emphasis on southeastern coyotes. Ecology and Evolution 9: 3389–3404.
Hody, J. W., & Kays, R. 2018. Mapping the expansion of coyotes (Canis latrans) across North and Central America. ZooKeys 759: 81.
Jeong, D. H., Jang, K. Yang, J. J., Choi, J. Y., Lim, S. H., Yeon, S. C., Shim, K. M., Kim, S. E., & Kang, S. S. 2021. Treatment of two Asiatic black bears (Ursus thibetanus) with severe injuries and their subsequent release into the wild: A case report. BMC Veterinary Research 17: 1–11.
Johnson, D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology 61: 65–71.
Kays, R., Crofoot, M. C., Jetz, W., & Wikelski, M. 2015. Terrestrial animal tracking as an eye on life and planet. Science 348: 1222.
Kilkenny C., Browne W. J., Cuthill I. C., Emerson M., & Altman D. G. 2010. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS Biology 8: e1000412.
Lamb, C., Smit, L., McLellan, B., Vander Vennen, L. M., & Proctor, M. 2022. Considerations for furbearer trapping regulations to prevent grizzly bear toe amputation and injury. Wildlife Society Bulletin 46: e1343.
Lindsjö, J., Fahlman, Å., & Törnqvist, E. 2016. Animal welfare from mouse to mouse – implementing the principles of the 3Rs in wildlife research. Journal of Wildlife Disease 52: S65–S77.
Lord, J., & Miller, E. A. 2019. Natural history and medical management of canids. In pages 313–325, Hernandez, S. M., Barron, H. W., Miller, E. A., Aguilar, R. F., and Yabsley, M. J., eds. Medical management of wildlife species: a guide for practitioners. John Wiley & Sons, Hoboken, NJ.
Lynn, W. S., Baker, L., Santiago-Ávila, F., & Stewart, K. L. Ethics, wellbeing, and wild lives. In pages 438–452, Fine, A. H., Mueller, M. K., Ng, Z. Y., Beck, A. M., and Peralta, J. M., eds. The Routledge International Handbook of Human-Animal Interactions and Anthrozoology. Routledge, New York, NY.
Lyons, A. J., & Getz, W. M. 2018. tlocoh: Time local convex hull home range and time use analysis. R package version 1.40.05. https://CRAN.R-project.org/package=tlocoh
Lyons, A. J., Turner, W. C., & Getz, W. M. 2013. Home range plus: a space-time characterization of movement over real landscapes. Movement Ecology 1: 1–14.
Manly, B. F. J., Lyman, L., McDonald, D., Thomas, L., McDonald, T. L., and Erickson, W. P. 2002. Resource selection by animals: statistical analysis and design for field studies, 2nd edition. Kluwar Academic, Boston, MA.
Mestre, F., Bragança, M. P., Nunes, A., & dos Santos, M. E. 2014. Satellite tracking of sea turtles released after prolonged captivity periods. Marine Biology Research 10: 996–1006.
Millspaugh, J. J., & Marzluff, M. 2001. Radio tracking and animal populations. Academic Press, San Diego, California.
Morin, D. J., & Kelly, M. J. 2017. The dynamic nature of territoriality, transience and biding in an exploited coyote population. Wildlife Biology 2017: wlb.00335.
Nájera, F., Uiterwaal, S. F., Crespo, E., Grande-Gómez, R., Sánchez, J. F., Mata-Huete, M., Palmer, J., Iturrarte, G., Peña, J., Munkhtsog, B., Munkhtsog, B., Poyarkov, A. D., Hernandez-Blanco, J. A., Alexandrov, D. Y., Galsadorj, N., & Deem, S. L. 2024. Insights into the spatial ecology of severly injured free-living felids: Iberian lynx, bobcat, and snow leopard. Ecology and Evolution 14: e11000.
Onderka, D. K., Skinner, D. L., & Todd, A. W. 1990. Injuries to coyotes and other species caused by four models of footholding devices. Wildlife Society Bulletin 18: 175–182.
Powell, R. A., & Proulx, G. 2003. Trapping and marking terrestrial mammals for research: Integrating ethics, performance criteria, techniques, and common sense. ILAR Journal 44: 259–276.
R Core Development Team. 2020. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Raine, A. F., Anderson, T., Vynne, M., Driskill, S., Raine, H., & Adams, J. 2020. Post-release survival of fallout Newell’s shearwater fledglings from a rescue and rehabilitation program on Kaua’I, Hawai’i. Endangered Species Research 43: 39–50.
Rio-Maior, H., Beja, P., Nakamura, M., Santos, N., Brandáo, R., Sargo, R., Dias, I., Silva, F., & Álvares, F. 2016. Rehabilitation and post-release monitoring of two wolves with severe injuries. Journal of Wildlife Management 80: 729–735.
Schweiger, A. K., Schütz, M., Anderwald, P., Schaepman, M. E., Kneubühler, M., Haller, R., & Risch, A. C. 2015. Foraging ecology of three sympatric ungulate species–behavioural and resource maps indicate differences between chamois, ibex and red deer. Movement Ecology 3: 1–12.
Sikes, R. S., and Animal Care and Use Committee of the American Society of Mammalogists. 2016. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. Journal of Mammalogy 97: 663–688.
Soulsbury, C. D., Gray, H. E., Smith, L. M., Braithwaite, V., Cotter, S. C., Elwood, R. W., Wilkinson, A., & Collins, L. M. 2020. The welfare and ethics of research involving wild animals: A primer. Methods in Ecology and Evolution 11: 1164–1181.
Sperry, C. C. 1939. Food habits of peg-leg coyotes. Journal of Mammalogy 20: 190–194.
Van Patter, L. E. 2022. Individual animal geographies for the more-than-human city: storying synanthropy and cynanthropy with urban coyotes. Nature and Space 5: 2216–2239.
Vogelnest, L. 2008. Veterinary considerations for the rescue, treatment, rehabilitation and release of wildlife. In pages 1–12, Medicine of Australian mammals. Eds L. Vogelnest and R. Woods. CSIRO Publishing, Collingwood, Victoria, Australia.
vonHoldt, B. M., Hinton, J. W., Shutt, A. C., Murphy, S., Karlin, M. L., Adams, J. R., Waits, L. P., and Brzeski, K. E. 2022. Reviving ghost alleles: Genetically admixed coyotes along the American Gulf Coast are critical for saving the endangered red wolf. Science Advances 8: eabn7731.
Ward, J. N., Hinton, J. W., Johannsen, K. L., Karlin, M. L., Miller, K. V., & Chamberlain, M. J. 2018. Home range size, vegetation density, and season influences prey use by coyotes (Canis latrans). PLoS ONE 13: e0203703.
Webster, S. C., Beasley, J. C., Hinton, J. W., & Chamberlain, M. J. 2022. Resident and transient coyotes exhibit differential patterns of movement behavior across heterogeneous landscapes in the southeastern United States. Ecology and Evolution 12: e8725.
Wilhelm, S. I., Schau, J. J., Schau, E., Dooley, S. M., Wiseman, D. L., & Hogan, H. A. 2013. Atlantic puffins are attracted to coastal communities in Eastern Newfoundland. Northeastern Naturalist 20: 624–630.
Willette, M., Rosenhagen, N., Buhl, G., Innis, C., and Boehm, J. 2023. Interrupted lives: Welfare considerations in wildlife rehabilitation. Animals 13: 1836.
Young, J., Schultz, J. T., Jolley, B., Baili, N., & Draper, J. P. 2022. Social learning of avoidance behaviors: Trap aversion in captive coyotes. USDA Wildlife Service, Staff Publications, 2587.
Accepted by 4 of 5 reviewers
Open Access
Peer-Reviewed
Creative Commons
Submitted: 20 December 2023
Accepted: 29 May 2024
Published: 03 July 2024
Funding Information: This study was funded through support received from the USFWS (F20AC11140-00) and the Wolf Conservation Center.
Conflicts of Interest: The authors declare no conflicts of interest.
© 2024 Hinton et al. The Stacks



