

Use of multi-modelling methods to inform conservation and reintroductions of pine marten Martes martes in Britain
* This article is part of a Special Issue from the Martes Working Group.
1 Vincent Wildlife Trust, Ledbury HR8 1EP, Herefordshire, UK.
JM: https://orcid.org/0000-0002-6919-120X
PW: https://orcid.org/0000-0002-1026-6596
2 United States Environmental Protection Agency, 200 SW 35th Street, Corvallis, Oregon, USA.
https://orcid.org/0000-0002-4331-825X
3 Computational Ecology Group Inc., Canmore, AB T1W 1P2, Canada.
SW: https://orcid.org/0000-0002-8542-7374
MacPherson, J., Wright, P., Schumaker, N., & Watkins, S. (2024). Use of multi-modelling methods to inform conservation and reintroductions of pine marten Martes martes in Britain. Stacks Journal: 24004. https://doi.org/10.60102/stacks-24004

Abstract photo. Two pine martens photographed in Scotland (© Robert Cruikshanks).
Abstract
The successful onset of recovery of the European pine marten (Martes martes) in some parts of Britain through range expansion and, more recently translocation for reintroductions, has resulted in a strong interest in reintroduction projects throughout the country. However, the geographic scope and conservation goals of these initiatives are often local and lack consideration of how they fit within the wider context of national-scale pine marten conservation. Here, we aim to maximize conservation benefit strategically at a national level by developing a simple, transparent, and transferable framework based on landscape modelling methods and spatially explicit population viability analyses. Our new methodology has been developed specifically to inform decisions involving the spatial targeting of pine marten conservation measures. We began by applying habitat suitability and connectivity modelling at a national scale. Then, we performed spatially explicit life history simulations to assess the natural recovery of the species. This information was used to identify regions of interest for future reintroductions, and we performed subsequent simulations to assess the viability of a reintroduced population within each region. From all the regions assessed, we identified two that should be prioritized for further consideration based on our analyses of habitat suitability, connectivity and the viability of reintroduced populations. While our framework can be used to identify and prioritize regions of conservation value generally, our focus here is on the biological considerations associated with identifying suitable landscapes for pine marten reintroduction.
Keywords: Circuitscape, conservation, HexSim, MaxEnt, Pine marten, reintroduction, translocations
Introduction
Conservation translocations and reintroductions are inherently complex, high-risk activities, and reviews of their outcomes have typically reported low rates of success (Wolf et al., 1996, Fischer & Lindenmayer, 2000, Seddon, 2010, Sainsbury & Vaughan‐Higgins, 2012, Weise et al., 2014, Berger‐Tal et al., 2020). Nonetheless, in circumstances where natural recovery or recolonization is unlikely, translocations may be necessary to re-establish a species following extirpation, ensure population viability and restore or expand geographic distribution.
Predictive models can play a key role in informing species reintroduction decision making (Lipsey et al., 2007). IUCN guidelines recommend that modelling be used to explore different scenarios and to devise an optimal strategy (IUCN/SSC, 2013). Time spent modelling the likely outcomes of translocation efforts is therefore disproportionately valuable, given the financial, welfare, and reputational costs of (often repeated) reintroduction failures. Habitat suitability models (HSMs) are commonly used to help inform such decisions by identifying regions with suitable resources that have the greatest potential to serve as contemporary reintroduction sites. However, HSMs alone cannot account for the population processes required for species persistence (Guisan et al., 2013). Using process models such as population viability analyses (PVA) in combination with HSMs can provide more information on the likely effects of conservation actions on long-term species persistence (Guisan et al., 2013). HSM-PVA hybrid models also have the potential to inform complex multi-endpoint strategies attempting to balance ecological benefits against economic costs and social concerns. One published example of such an approach, used to assess critical habitats for Ord’s kangaroo rat (Dipodomys ordii) in Alberta, Canada (Heinrichs et al., 2010), showed that 39% of habitat predicted to be suitable for this species was unlikely to contribute to population viability. This and other examples illustrate that, when forecasting species persistence is a priority concern, insights gained from the coupling of HSMs and spatial PVA models can prove uniquely valuable (Larson et al., 2004, Andersen et al., 2017).
The pine marten (Martes martes) is a medium-sized mustelid associated predominantly with forested habitat (Mitchell-Jones et al., 1999). The species occurs throughout most of continental Europe, Asia Minor, northern Iraq and Iran, the Caucasus and in westernmost parts of Asian Russia. Pine martens were once common and widespread throughout Britain (Maroo & Yalden, 2000), but during the 19th and early 20th centuries the population suffered severe declines in numbers and distribution. This trajectory resulted from increases in predator control (Langley & Yalden, 1977, Tapper, 1992), coupled with the ongoing loss and fragmentation of woodland habitat. By the beginning of the 20th century, the pine marten was almost exclusively restricted to the north-west highlands in Scotland, and much smaller habitat remnants in the remote uplands of northern England and Wales (Langley & Yalden, 1977). Subsequent increases in afforestation and legal protection since the 1980s have allowed the pine marten to recover some of its former range in Scotland, but not elsewhere in Britain. In Wales, the population was thought to be functionally extinct. Thus, following a feasibility study (MacPherson et al., 2014), a total of 51 pine martens were translocated to Wales from Scotland between 2015 and 2017. This was followed (in 2019 and 2021) by a reintroduction of 35 individuals to the Forest of Dean in Gloucestershire, on the English/Welsh border. The successful establishment of both of these populations has led to an increasing number of reintroduction efforts being proposed throughout Britain, in part because recent studies in Ireland (Sheehy & Lawton, 2014) and Scotland (Sheehy et al., 2018) suggest that pine martens may have a negative impact on the invasive grey squirrel (Sciurus carolinensis). Local organizations and partnerships throughout Britain are particularly interested in using reintroduced pine marten populations as a cost-effective means for controlling grey squirrels to the benefit of commercial forestry, as well as native red squirrels (Sciurus vulgaris). Yet, in spite of their goal to reintroduce the species, these local initiatives are not typically developed strategically with national-scale pine marten conservation in mind. Reintroductions planned at a local scale may actually hamper the species’ long-term recovery because the source population for translocations is limited, and wide-ranging carnivores, like pine martens, require landscapes with sufficiently large blocks of well-connected habitat to provide enough contiguous home ranges for a viable population. For this reason, a strategic approach is needed.
In this study, we aimed to develop a simple and transparent framework, based on combining widely used modelling methods. These are usually applied in isolation, however by combining them in this way they can inform the spatial targeting of pine marten conservation measures and help realize the maximum conservation benefit at a national level.
Methods and Materials
Framework
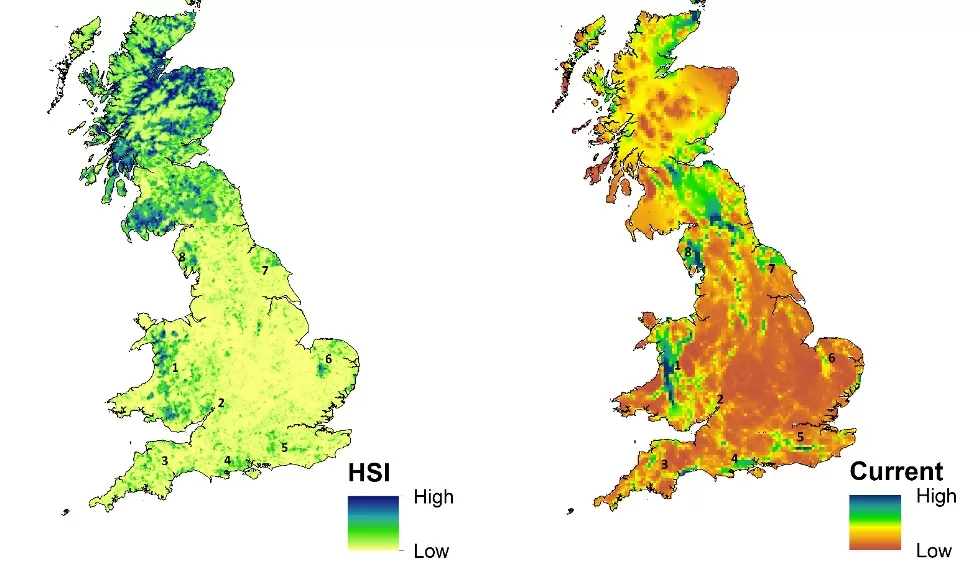
We developed a framework to help guide the spatial targeting of reintroductions, using modelling methods based on data and knowledge of pine marten ecology and distribution (Fig. 1). Our approach initially focuses on identifying habitat suitability and connectivity, and the species’ potential for natural (unaided) range recovery. Next, we evaluate the suitability of candidate translocation recipient regions to host a viable population. We applied the second part of the framework to six regions in Britain (regions 3-8; Fig. 2) to demonstrate its use and assess their potential for pine marten recovery or restoration.
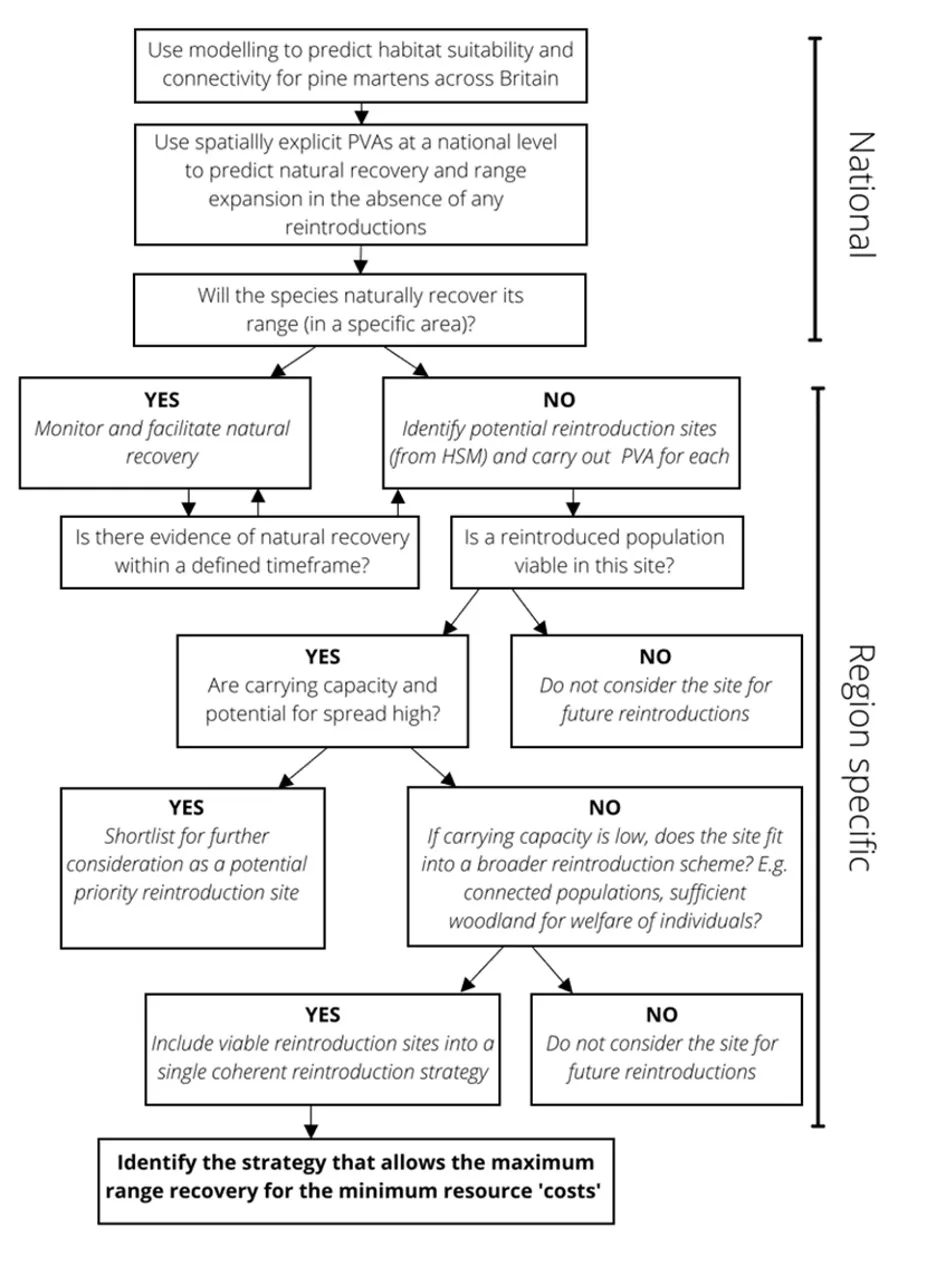
Habitat suitability modelling
We used MaxEnt (Phillips et al., 2006), a presence-only Habitat suitability modelling (HSM) approach, to predict suitability of pine marten habitat throughout Britain and identify potential reintroduction sites. All analyses were carried out using R (v. 3.5.3; R Core Team, 2021) in R Studio (v.1.2.5042; RStudio Team). Pine marten presence location records were available from Britain, Ireland, Northern Ireland and the Netherlands so we ran multiple models using different combinations of these datasets and compared them with Akaike Information Criteria (AIC) (Bozdogan, 1987). We measured the correlation between their outputs using Schoener’s D to assess the consistency in our predictions (Warren et al., 2010). (Results from the full suite of models are reported in MacPherson and Wright, 2021).
Having explored the alternatives, we utilized models based on the MaxEnt model trained on records from Britain. This had the lowest AIC and the highest AUC (Area Under the Curve), suggesting the best fit. Ecologically, this also made sense, as the suite of potential competitors and prey for pine martens is different in continental Europe, and thus there may be subtle differences in the species’ habitat utilization and ecology in the Netherlands. The model built on data from Britain resulted in a slightly more conservative map of predicted habitat suitability than that incorporating data from the Netherlands. However, in the context of the decision framework, the consequences of overestimating habitat suitability would be far worse than those of underestimating it.
Presence records consisted of pine marten scats, confirmed by DNA testing, which had been collected during two surveys taking place between 2012-2013 in Scotland (Croose et al., 2013, Croose et al., 2014). Both surveys were based on 1-3 transects of 1-1.5 km in length, located within each 10 km grid square (hectad). Experienced surveyors walked transects located along forest tracks or paths in wooded habitat searching for pine marten scats. Each transect was walked only once between May and September. Additional camera trap records collected by Vincent Wildlife Trust (VWT) from 2017 to 2020 during the ‘Back From The Brink’ project in northern England were also included in the analysis (https://www.vwt.org.uk/projects-all/back-from-the-brink/ ). We also obtained pine marten records from across Scotland that were stored within the GBIF database (GBIF.ORG, 2020). We only used validated GBIF records with a precision of at least 1000m, dating from 2005 onwards. For each model, we filtered records to retain a single record per grid square.
We used a suite of environmental variables (shown in table 1) derived from Wright et al. (2020) and available at two candidate scales – 1 km and 3 km squares (MacPherson and Wright, 2021). These comprised information on habitat cover, but also provided information on the impact of road density and traffic. The optimal scale was identified for each predictor by creating univariate models using default settings with threshold features disabled (Bellamy et al., 2020, Wright et al., 2020, Hijmans et al., 2017). The scale with the highest training gain measure was then selected, which can be interpreted as the likelihood of the presence points (Merow et al., 2013, Bellamy et al., 2020).
For each model, we removed highly correlated variables using the ‘vifstep’ stepwise function of the ‘usdm’ package (Naimi et al., 2014) and a conservative VIF threshold of three (Zuur et al., 2010). We used the package ‘ENMeval’ (Muscarella et al., 2014) to identify the optimal MaxEnt model settings. We tested combinations of feature types (L, linear; H, hinge; Q; quadratic; P, product) and disabled threshold features to reduce overfitting. We varied the regularization multiplier in steps of 0.5, from 0.5 to 4. Then, we developed a final model using the optimal settings to produce model predictions.
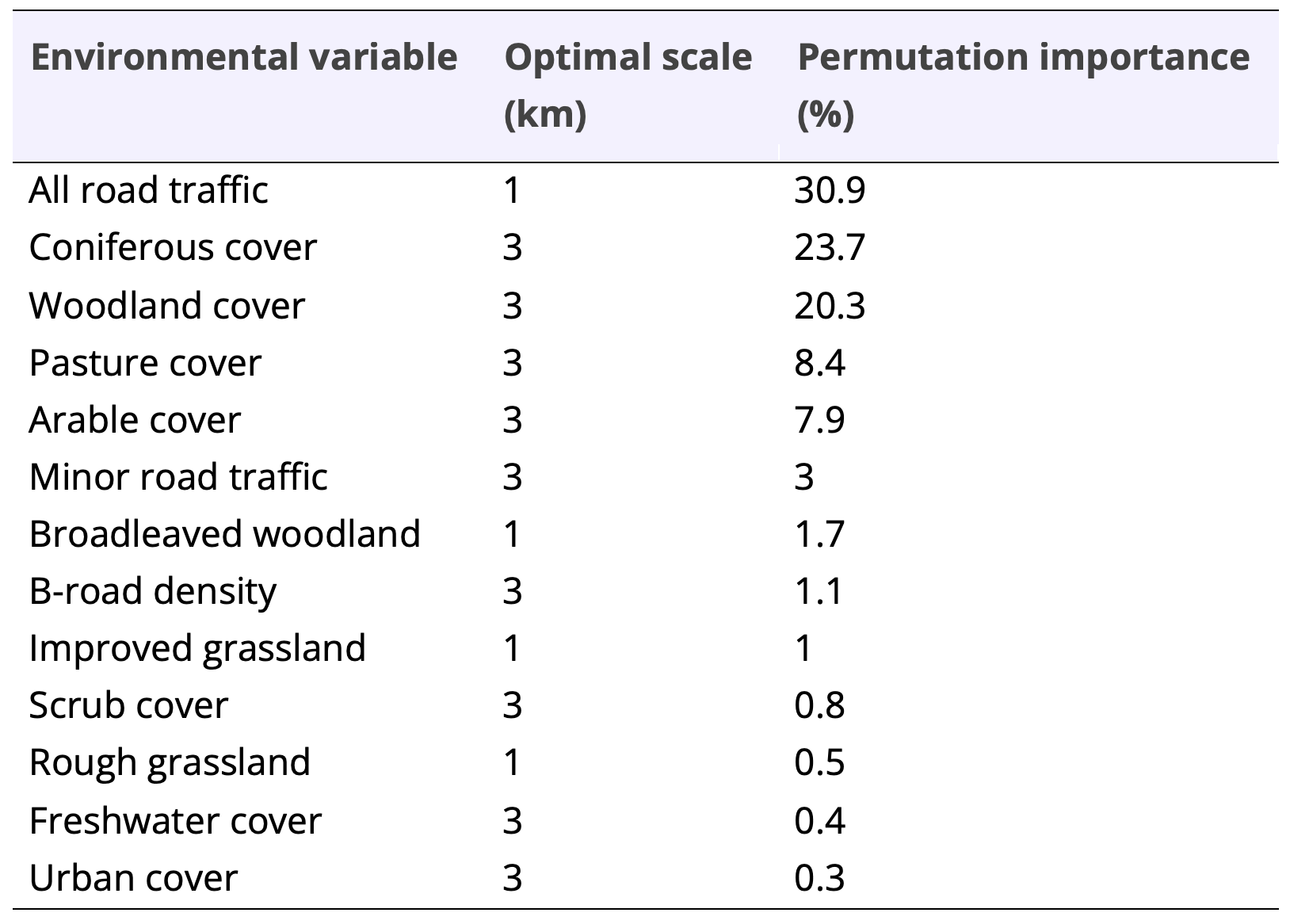
Table 1. Environmental predictor contribution values.
Connectivity modelling
Here, we used resistance-surface-based connectivity modelling where the values of each grid cell of the resistance surface were indicative of the amount of movement possible. The resistance surface ranged from 1 to 100 and was based on our pine marten HSM. To transform the habitat suitability values into a resistance surface, we used a negative exponential function allowing the dispersal of pine marten through unsuitable habitat (Trainor et al., 2013, Mateo-Sánchez et al., 2015, Keeley et al., 2016):
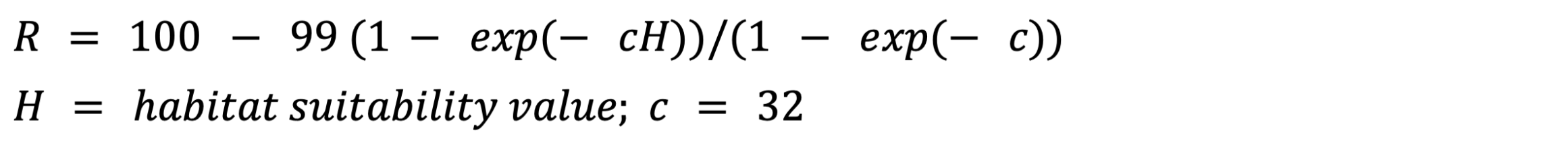
Finally, we used Circuitscape v4.0.5, a software which calculates all possible pathways between nodes in a landscape using the circuit theory (McRae & Nürnberger, 2006), to perform “pairwise” calculations of the amount of current flowing between a combination of nodes. The current flowing is representative of the pathways that pine martens are likely to use especially for the dispersal of individuals. To place nodes, we followed a similar approach to Koen et al. (2014) and we created a 120 km buffer zone around Britain with a resistance score of 100. We randomly selected 50 points (focal nodes) within the buffer zone. By placing focal nodes outside of the study area, we removed most of the biases introduced by node placement which gives a better representation of possible animal movement when there is not a specific need to place nodes inside the landscape. The cumulative current map resulting from all pairwise tests was then used as the final Circuitscape output once the buffer zone was removed to identify areas of high connectivity in Britain.
HexSim models
HexSim (Schumaker & Brookes, 2018) is a life history simulator used for building population viability models that is ideal for incorporating species’ interactions and responses to disturbance. HexSim models are spatially explicit and individual-based, and individuals can be assigned dynamic life history traits. In our simulations, we first investigated the spread of pine martens from the Scottish borders across Britain, over a period of more than 25 years, with and without reintroductions into central Wales and the Forest of Dean. Our initial simulated translocations were designed to replicate actual translocations consisting of the releases of 20 pine martens (10 males and 10 females) into central Wales each year over a period of three consecutive years, followed by two years of reintroductions into the Forest of Dean (Fig. 2 – sites 1-2). The initial simulated population was set to 3,700 individuals which equates to the most recent estimates by Mathews et al. (2018). At simulation year 6, the first year of translocations in central Wales took place, followed by the Forest of Dean.
We subsequently investigated the viability of reintroduced pine marten populations at regions of conservation interest (Fig. 2 – sites 3-8) by performing similar reintroduction scenarios over a 50 year period, assuming a closed population with no emigration from or immigration into the study area, to assess the reintroduction success in specific areas.
Our HexSim models were designed to replicate the principal events occurring within the pine marten life cycle. Population and life history parameters were derived from Powell et al. (2012). As is the case in the wild, our simulated pine martens were territorial, maintaining exclusive core areas, and would not tolerate territory overlap with individuals of the same sex. Simulated males had larger home range sizes than females and could overlap with more than one female. Territorial martens acquired resources from their home ranges, and those lacking territories were left to make use of any remaining unclaimed resources. We used the maps generated by our HSM as a habitat map in our simulation models; it was this map that indicated resource availability.
The simulated marten life cycle events proceeded in order, as follows: (1) At the beginning of each year all martens got older, and juveniles and subadults incremented their stage classes. Thus, at this moment in time, the population will consist exclusively of subadults and adults. In select years, reintroductions were performed. All reintroduced females were assumed to be pregnant. (2) Subadults dispersed, and adults, followed by subadults, prospected for territories, which ensured that adults had an advantage in resource acquisition. Pine martens (real and simulated) will not abandon a territory, thus the only adults who prospected were those who previously failed to obtain a territory. Prospecting martens were allowed to make short-distance excursions as necessary to move between nearby resource patches (maximum distance of 50 hexagons, equivalent to approximately 27km) . (3) Males and females formed reproductive pairs based on proximity, but this also involved a scramble competition. The simulated martens used a polygamous mating system, and only paired females were allowed to reproduce. (4) All simulated martens were subject to annual mortality, with mortality rates varying based on stage class and resource availability. Senescence was imposed by setting the survival rate of 8-year-old martens to zero.
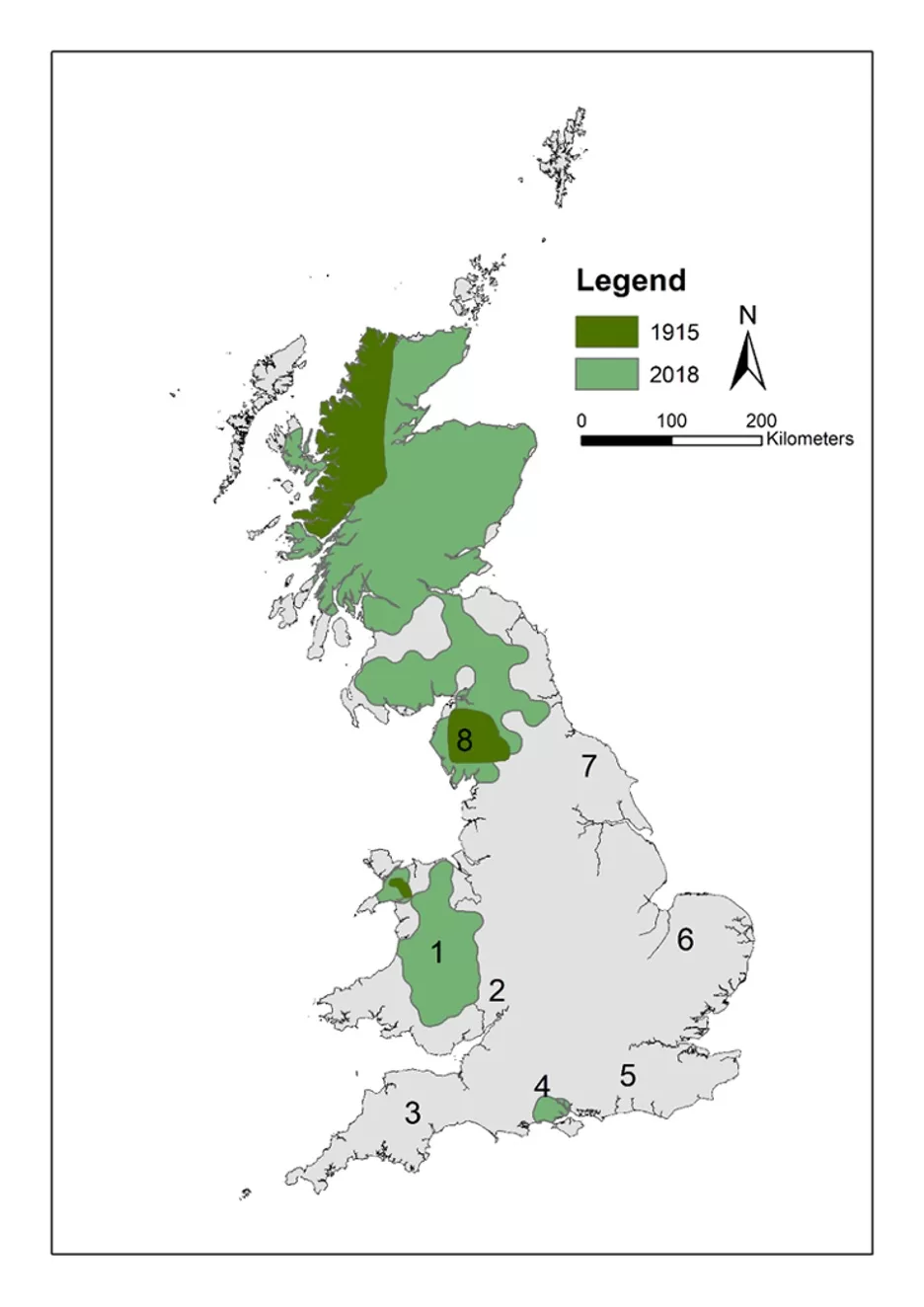
Figure 2. Presumed pine marten distributional range in Britain in 2018 as reported by Mathews et al. (2018) and 1915 (Langley & Yalden, 1977). Translocations took place in 2015-17 in region 1 (1. Mid-Wales) and in 2019-21 in region 2 (2. Forest of Dean, Gloucestershire). Regions 3-8 are regions considered in detail in this study (3. South West England, 4. Hampshire, 5. Southeast England, 6. East Anglia, 7. North Yorkshire, 8. South Cumbria). It should be noted that, as a consequence of the methods used, Mathews et al (2018) likely over-estimated the distributional range of pine martens in region 8 where the very few recent records provide no evidence of an established breeding population.
Results
Habitat suitability and connectivity modelling
Our final habitat suitability model (LQHPT-2) used 1,808 pine marten records and had an AUC of 0.93 (average AUC = 0.91 ± 0.002). Pine marten presence was strongly associated with reduced road traffic (1 km scale) and high coniferous woodland (3 km scale) and woodland cover (3 km scale; Shown in Table 1).
The habitat suitability and connectivity modelling suggest that the majority of remaining well-connected highly suitable habitat for the species is found in the north and west of Britain (Fig. 3).
Population modelling – National recovery
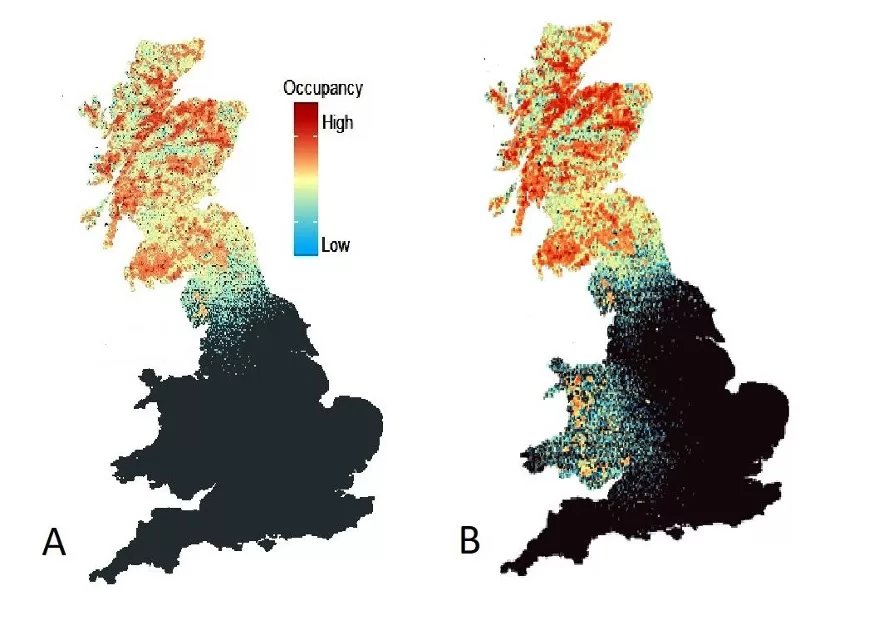
Results from our PVA model (Fig. 4) suggest that, within 25 years, the pine marten population in Scotland is likely to continue spreading south and become established in several of the northern counties of England, including Cumbria, Northumberland and Durham. Over this timescale it is also probable that pine martens will have begun to expand into Lancashire and North Yorkshire. Without intervention, however, the pine marten population is unlikely to show any natural expansion further south than this in England.
Simulation results incorporating translocations that have already taken place suggest that the re-established population in Wales is robust enough to support significant and rapid range expansion (Fig. 4), and will be contiguous with the reintroduction to the Forest of Dean on the English-Welsh border within 25 years. However, we did not observe any natural recolonization of suitable habitats in south-west England or in the east of the country within this timeframe (region 3-6).
Population modelling – Regional reintroductions
The results from our simulated regional reintroductions varied as follows (see Fig. 2):
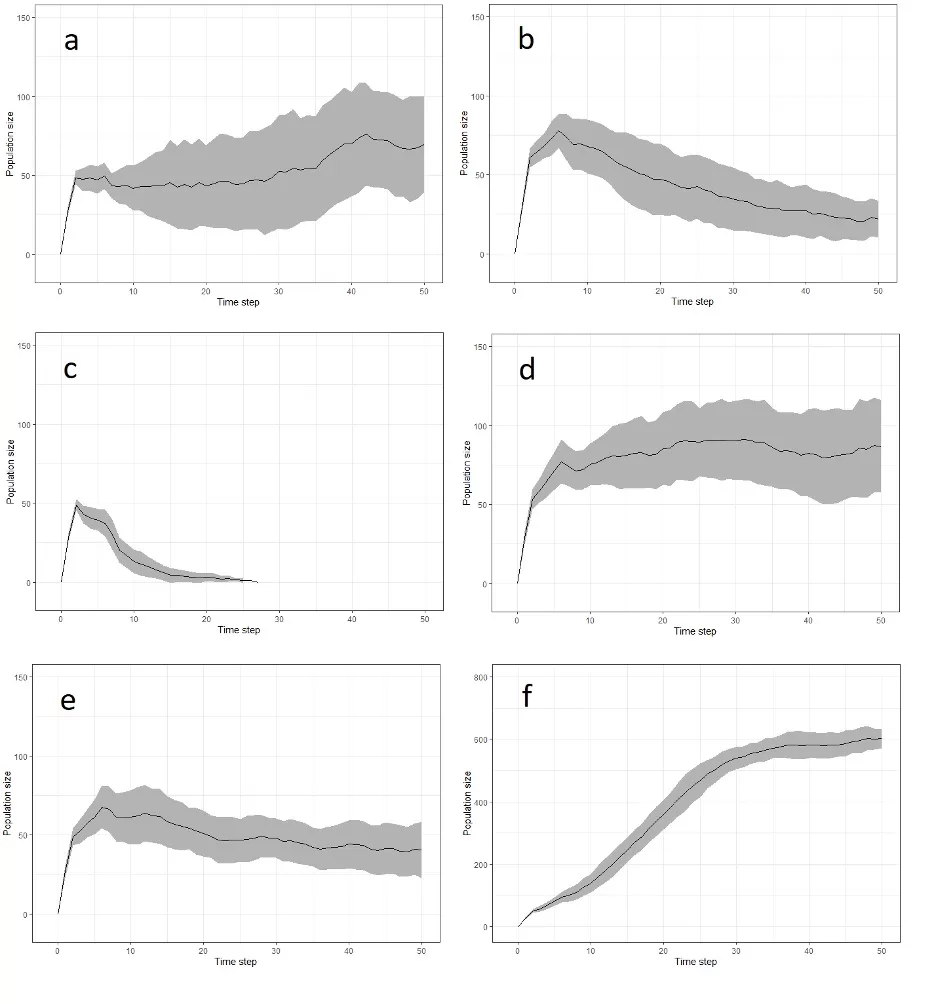
Region 3 South West England – Suitable habitat was dispersed throughout the region, and connectivity was high across the north and south. The national-scale HexSim models predicted that this region will not be recolonized by the expanding population from regions 1 and 2, despite their relative proximity. The simulations suggest that reintroductions will likely result in a viable population (Fig. 5a).
Region 4 Hampshire – Both suitable habitat and connectivity were present in this region. However, our simulated reintroduction of pine martens into this region produced a local population that, while declining, will persist for 50 years, albeit at very low levels (Fig. 5b). The national-scale HexSim models predicted that this region will not be recolonized by the expanding population.
Region 5 Southeast England – Our analyses suggest that, despite there being suitable habitat in this region, other factors (specifically very high road and traffic density and concentration of human population) make this region unsuitable for pine marten reintroductions. In the establishment phase, our PVA suggests that mortality would exceed reproduction and recruitment (Fig. 5c).
Region 6 East Anglia – Habitat suitability was high but connectivity across this region is very low, despite the presence of considerable suitable habitat. As a consequence, pine martens in our simulations had limited ability to disperse away from the release area once the carrying capacity had been exceeded (Fig. 5d).
Region 7 North Yorkshire – High habitat suitability and connectivity was found in this region, but it is fairly isolated from other areas to the north. Our simulation model results (Fig. 4) indicate that this region is unlikely to be naturally colonized from an expanding Scottish population. Though it persisted for 50 years, the simulated reintroduced population here declined slowly over time (Fig. 5e). Ultimately, the population fell below the number of animals released, and as such became highly vulnerable to stochastic events.
Region 8 South Cumbria – Our HSM model suggests that there is a large amount of suitable habitat available to pine martens here. There have been a small number of intermittent records from this region over a number of years but no evidence to date of an established population. Our national-scale HexSim model predicted that this region will be naturally recolonized by pine martens within 25 years, so the recommendation would be to monitor and assess. However, in the event that recolonisation is not observed within a reasonable timeframe, we ran our PVA model which indicates that reintroductions would lead to a viable population (Fig. 5f).
Discussion
The prioritization of pine marten reintroduction regions in Britain is key to maximize the conservation benefit of the species at a national level. Our results suggest that south-west England (region 3), and potentially south Cumbria (region 8), merit further consideration as potential pine marten reintroduction regions. These regions may be suitable for reintroductions contingent on the results of appropriate disease risk analysis, as well as risk assessments for other species and habitats and minimal conflict with other land users. Implementing such a policy would also depend on sufficient resources being secured for reintroduction and subsequent long-term monitoring and engagement.
Their context within anthropogenically altered landscapes is fundamental to the success of carnivore reintroductions. In south-east England and Hampshire/New Forest (regions 4 and 5), the high density of roads, traffic and other infrastructure may pose too high a threat to a founder pine marten population in comparison to one at carrying capacity (Deredec & Courchamp, 2007). Though this area does have suitable habitat for pine martens, the dense road network and relatively high proportion of land where gamebirds are reared and released for commercial shooting may mean increased risks of accidental mortality as a result of lawful predator control.
Similar barriers are present further north (regions 7 and 8) as conurbations, and associated road networks surrounding these regions are expected to impede natural colonization. It is possible that a small but constant influx of individuals dispersing from the north might make a reintroduced population of pine martens more resilient in region 7. In region 8 (Cumbria), recolonisation is predicted therefore natural recovery might be prioritized here over translocations, and this area should be monitored and facilitated where possible. However, it must also be noted that the expanding population in southern Scotland may derive from a relatively small number of animals that were translocated to in the early 1980s (Shaw & Livingstone, 1992) with consequently low levels of genetic diversity, which may serve to lower the effective rate of range expansion in the Scottish borders (Croose et al., 2013). Some rehabilitated martens have also subsequently been released here but in numbers far too small to effect any genetic rescue. It was not possible to include this or to vary the rate of population expansion across the current range within our HexSim model. Therefore, we suggest that if there is no evidence of significant natural recolonization in Cumbria, then the potential for reintroductions should be explored. This would be an effective way of accelerating the rate of spread of pine martens in northern England.
Considerable effort should be put into expanding and improving existing habitat in terms of size, quality and connectivity, providing good natural and artificially enhanced dispersal corridors. A case in point is East Anglia (region 6 here), where we found one area of high habitat suitability but with little or no connectivity to anything beyond. As a consequence, pine martens would have no way of dispersing away once the carrying capacity is exceeded, resulting in an isolated ‘island’ population with attendant issues of genetic management. The potential for increasing habitat connectivity and linkages between established pine marten populations and the regions considered in this study should be explored to create dispersal corridor routes, as well as possible road crossings/green bridges/underpasses to increase landscape permeability for pine martens and other wildlife. Although the effectiveness of these is highly dependent on context (Smith et al., 2015, Kusak et al., 2009), there is some evidence that martens will use them (Georgii et al., 2011).
The framework presented here was developed with the aim of informing decisions around spatial targeting of pine marten conservation measures. However, it should not be the sole method used for making decisions regarding if and where reintroductions should take place. Reintroductions must consider social, as well as biological feasibility in the context of the species, habitats and landscapes where they are being considered. Local support and stakeholder participation in the decision process is vital for the long-term establishment (IUCN/SSC, 2013), particularly of a carnivore reintroduction.
It is also essential to ensure that harvesting of donor populations for reintroductions is sustainable. In addition to any impact on absolute numbers, harvesting donor populations can potentially cause a genetic bottleneck leading to inbreeding and loss of genetic diversity, and as a result may have hidden consequences (O’Reilly et al., 2024). Therefore, the size and genetic diversity of potential donor populations are central points to consider when assessing impacts of removals on both genetic diversity and population persistence (MacPherson et al., 2014, IUCN/SSC, 2013). It is essential to monitor the impacts of translocations to ensure that harvesting of donor populations for reintroductions is sustainable (MacPherson et al., 2020) and to safeguard viable populations of pine martens throughout their range in Britain.
Conclusion
Habitat suitability and connectivity modelling showed that the majority of well-connected, highly suitable habitat for pine martens is in the north and west of Britain. Translocations of pine martens have already taken place for reintroductions into Wales, with the largest contiguous region of highly suitable habitat outside of Scotland, and subsequently to the Forest of Dean in England. Our structured decision-making process for prioritizing suitable regions for further investigation identified two regions that should be prioritized for further consideration based on landscape modelling methods and spatially explicit population viability analyses. This framework, however, is focused on the biological considerations associated with identifying potentially suitable landscapes for pine martens. It must be stressed that any conservation translocation also needs to consider and address relevant socio-economic and regulatory considerations.
Acknowledgments
We would like to thank Robert Raynor, Kat Walsh, Claire Howe and Steve Carter for their invaluable help and comments. The information in this document has been funded in part by the U.S. Environmental Protection Agency. It has been subjected to review by the Center for Public Health and Ecological Assessment and approved for publication. Approval does not signify that the contents reflect the views of the Agency, nor does mention of trade names or commercial products constitute endorsement or recommendation for use.
Author Contributions
Jenny MacPherson: Conceived the ideas and designed the methods, Led the writing
Patrick Wright: Conceived the ideas and designed the methods, Carried out the analyses
Nathan Schumaker: Assisted with analyses
Sydney Watson: Assisted with analyses
All authors contributed to the drafts and gave final approval for publication.
Data Availability
Data from this research are subject to data-sharing agreements with collaborators. We are happy to provide data from this research upon request. Code for these analyses can be found at
https://cran.r-project.org/web/packages/maxnet/index.html;
https://cfpub.epa.gov/si/si_public_record_report.cfm?Lab=NHEERL&dirEntryId=339689;
https://docs.circuitscape.org/Circuitscape.jl/latest/
Transparent Peer Review
Results from the Transparent Peer Review can be found here.
Recommended Citation
MacPherson, J., Wright, P., Schumaker, N., & Watkins, S. (2024). Use of multi-modelling methods to inform conservation and reintroductions of pine marten Martes martes in Britain. Stacks Journal: 24004. https://doi.org/10.60102/stacks-24004
References
Andersen, L. H., Sunde, P., Pellegrino, I., Loeschcke, V. & Pertoldi, C. 2017. Using population viability analysis, genomics, and habitat suitability to forecast future population patterns of Little Owl Athene noctua across Europe. Ecology and evolution, 7, 10987-11001.
Bellamy, C., Boughey, K., Hawkins, C., Reveley, S., Spake, R., Williams, C. & Altringham, J. 2020. A sequential multi-level framework to improve habitat suitability modelling. Landscape Ecology, 1-20.
Berger‐Tal, O., Blumstein, D. & Swaisgood, R. R. 2020. Conservation translocations: a review of common difficulties and promising directions. Animal Conservation, 23, 121-131.
Bozdogan, H. 1987. Model selection and Akaike’s information criterion (AIC): The general theory and its analytical extensions. Psychometrika, 52, 345-370.
Croose, E., Birks, J. & Schofield, H. W. 2013. Expansion zone survey of pine marten (Martes martes) distribution in Scotland. Scottish Natural Heritage.
Croose, E., et al. (2014). Distribution of the pine marten (Martes martes) in southern Scotland in 2013, Scottish Natural Heritage Commisioned Report No. 740.
Deredec, A. & Courchamp, F. 2007. Importance of the Allee effect for reintroductions. Ecoscience, 14, 440-451.
Fischer, J. & Lindenmayer, D. B. 2000. An assessment of the published results of animal relocations. Biological Conservation, 96, 1-11.
GBIF.ORG. 2020. GBIF Occurrence Download [Online]. Accessed from R via rgbif (https://github.com/ropensci/rgbif) Available: https://doi.org/10.15468/dl.zez7ee [Accessed 2020-05-29].
Georgii, B., Keller, V., Pfister, H. P., Reck, H., Peters-Ostenberg, E., Henneberg, M., Herrmann, M., Mueller-Stiess, H. & Bach, L. 2011. Use of wildlife passages by invertebrate and vertebrate species. Wildlife passages in Germany, 2011, 1-27.
Guisan, A., Tingley, R., Baumgartner, J. B., Naujokaitis‐Lewis, I., Sutcliffe, P. R., Tulloch, A. I., Regan, T. J., Brotons, L., McDonald‐Madden, E. & Mantyka‐Pringle, C. 2013. Predicting species distributions for conservation decisions. Ecology letters, 16, 1424-1435.
Heinrichs, J. A., Bender, D. J., Gummer, D. L. & Schumaker, N. H. 2010. Assessing critical habitat: evaluating the relative contribution of habitats to population persistence. Biological Conservation, 143, 2229-2237.
Hijmans, R. J., Phillips, S., Leathwick, J. & Elith, J. 2017. Package ‘dismo’. Circles, 9, 1-68.
IUCN/SSC 2013. Guidelines for reintroductions and other conservation translocations. Version 1.0. IUCN Species Survival Commission Gland, Switzerland.
Keeley, A. T., Beier, P. & Gagnon, J. W. 2016. Estimating landscape resistance from habitat suitability: effects of data source and nonlinearities. Landscape ecology, 31, 2151-2162.
Koen, E. L., Bowman, J., Sadowski, C. & Walpole, A. A. 2014. Landscape connectivity for wildlife: development and validation of multispecies linkage maps. Methods in Ecology and Evolution, 5, 626-633.
Kusak, J., Huber, D., Gomerčić, T., Schwaderer, G. & Gužvica, G. 2009. The permeability of highway in Gorski kotar (Croatia) for large mammals. European Journal of Wildlife Research, 55, 7-21.
Langley, P. & Yalden, D. 1977. The decline of the rarer carnivores in Great Britain during the nineteenth century. Mammal Review, 7, 95-116.
Larson, M. A., Thompson Iii, F. R., Millspaugh, J. J., Dijak, W. D. & Shifley, S. R. 2004. Linking population viability, habitat suitability, and landscape simulation models for conservation planning. Ecological Modelling, 180, 103-118.
Lipsey, M. K., Child, M. F., Seddon, P. J., Armstrong, D. P. & Maloney, R. F. 2007. Combining the fields of reintroduction biology and restoration ecology. Conservation Biology, 1387-1390.
MacPherson, J., Croose, E., Bavin, D., O’Mahony, D., Somper, J. & Buttriss, N. 2014. Feasibility assessment for reinforcing pine marten numbers in England and Wales. Ledbury, UK: Vincent Wildlife Trust.
Macpherson, J., Croose, E., Powell, C., Carter, S. & O’reilly, C. 2020. Preliminary work towards a sustainable harvesting model of pine martens in Scotland for translocations. Ledbury, Herefordshire: Vincent Wildlife Trust.
MacPherson, J. & Wright, P. 2021. A Long-term Strategic Recovery Plan for Pine Marten Martes Martes in Britain. Ledbury, Herefordshire: Vincent Wildlife Trust.
Maroo, S. & Yalden, D. 2000. The mesolithic mammal fauna of Great Britain. Mammal Review, 30, 243-248.
Mateo-Sánchez, M. C., Balkenhol, N., Cushman, S., Pérez, T., Domínguez, A. & Saura, S. 2015. A comparative framework to infer landscape effects on population genetic structure: are habitat suitability models effective in explaining gene flow? Landscape Ecology, 30, 1405-1420.
McRae, B. H. & Nürnberger, B. 2006. Isolation by resistance. Evolution, 60, 1551-1561.
Merow, C., Smith, M. J. & Silander Jr, J. A. 2013. A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography, 36, 1058-1069.
Mitchell-Jones, A. J., Amori, G., Bogdanowicz, W., Krystufek, B., Reijnders, P., Spitzenberger, F., Stubbe, M., Thissen, J., Vohralik, V. & Zima, J. 1999. The atlas of European mammals, Academic Press London.
Muscarella, R., Galante, P. J., Soley‐Guardia, M., Boria, R. A., Kass, J. M., Uriarte, M. & Anderson, R. P. 2014. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods in Ecology and Evolution, 5, 1198-1205.
Naimi, B., Hamm, N. A., Groen, T. A., Skidmore, A. K. & Toxopeus, A. G. 2014. Where is positional uncertainty a problem for species distribution modelling? Ecography, 37, 191-203.
O’Rreilly, C., Sheehy, E., Macpherson, J., Birks, J., Martin, J., Croose, E., Fletcher, K., Lambin, X., Curran, T. & Synnott, R. 2024. Genetic Lag in a Demographically Recovering Carnivore: The Case of the British Pine Marten (Martes martes).
Phillips, S., Anderson, R. P. & Schapire, R. 2006. Maximum entropy modeling of species geographic distributions. Ecological Modeling, 190, 231-259. Phillips, S.(2006). A brief tutorial on Maxent.
Powell, R. A., Lewis, J. C., Slough, B. G., Brainerd, S. M., Jordan, N. R., Abramov, A. V., Monakhov, V., Zollner, P. A. & Murakami, T. 2012. 6. Evaluating Translocations of Martens, Sables, and Fishers: Testing Model Predictions with Field Data. Biology and Conservation of Martens, Sables, and Fishers. Cornell University Press.
R Core Team 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing,. Vienna, Austria.
RStudio Team RStudio: Integrated Development for R. Boston, MA. 2015. There is no corresponding record for this reference.[Google Scholar].
Sainsbury, A. W. & Vaughan‐Higgins, R. J. 2012. Analyzing disease risks associated with translocations. Conservation Biology, 26, 442-452.
Schumaker, N. H. & Brookes, A. 2018. HexSim: a modeling environment for ecology and conservation. Landscape Ecology, 33, 197-211.
Seddon, P. J. 2010. From reintroduction to assisted colonization: moving along the conservation translocation spectrum. Restoration Ecology, 18, 796-802.
Shaw, G. & Livingstone, J. 1992. The pine marten: its reintroduction and subsequent history in the Galloway Forest Park. Transactions of the Dumfries and Galloway Natural History and Antiquarian Society, third Series, 67, 1-7.
Sheehy, E. & Lawton, C. 2014. Population crash in an invasive species following the recovery of a native predator: the case of the American grey squirrel and the European pine marten in Ireland. Biodiversity and Conservation, 23, 753-774.
Sheehy, E., Sutherland, C., O’Reilly, C. & Lambin, X. 2018. The enemy of my enemy is my friend: native pine marten recovery reverses the decline of the red squirrel by suppressing grey squirrel populations. Proceedings of the Royal Society B: Biological Sciences, 285, 20172603.
Smith, D. J., Van Der Ree, R. & Rosell, C. 2015. Wildlife crossing structures: an effective strategy to restore or maintain wildlife connectivity across roads. Handbook of road ecology, 172-183.
Tapper, S. 1992. Game heritage: an ecological review from shooting and gamekeeping records, Game Conservancy Fordingbridge, UK.
Trainor, A. M., Walters, J. R., Morris, W. F., Sexton, J. & Moody, A. 2013. Empirical estimation of dispersal resistance surfaces: a case study with red-cockaded woodpeckers. Landscape ecology, 28, 755-767.
Warren, D. L., Glor, R. E. & Turelli, M. 2010. ENMTools: a toolbox for comparative studies of environmental niche models. Ecography, 33, 607-611.
Weise, F. J., Stratford, K. J. & van Vuuren, R. J. 2014. Financial costs of large carnivore translocations–accounting for conservation. PLoS One, 9, e105042.
Wolf, C. M., Griffith, B., Reed, C. & Temple, S. A. 1996. Avian and mammalian translocations: update and reanalysis of 1987 survey data. Conservation biology, 10, 1142-1154.
Wright, P. G., Coomber, F. G., Bellamy, C. C., Perkins, S. E. & Mathews, F. 2020. Predicting hedgehog mortality risks on British roads using habitat suitability modelling. PeerJ, 7, e8154.
Zuur, A. F., Ieno, E. N. & Elphick, C. S. 2010. A protocol for data exploration to avoid common statistical problems. Methods in ecology and evolution, 1, 3-14.
Accepted by 4 of 4 reviewers
Open Access
Peer-Reviewed
Creative Commons
Accepted: 28 August 2024
Published: 08 November 2024
Funding Information: This research was funded by Vincent Wildlife Trust, NatureScot and Natural England.
Conflicts of Interest: The authors declare no conflicts of interest.
© 2024 MacPherson et al. Stacks Journal



